
Согласованное взаимодействие нервных центров для обеспечения какого либо процесса это
Обновлено: 30.06.2024
4) функциональная система, ее задача – реализация определенных функций.
Принципы обработки информации
Изучение мозга – это фактически изучение того, как он обрабатывает информацию. Современная наука представляет себе этот процесс в самом общем виде, и выглядит это следующим образом:
– обработка происходит в форме последовательных этапов, каждый этап реализуется определенным нервным центром;
– на каждом из этапов выполняется уникальная операция;
– система функционирует по принципу отрицательной обратной связи: решение нервным центром задачи прерывает его активность.
Такой алгоритм обработки информации возможен благодаря тому, что нервные центры находятся в иерархическом соподчинении. Иерархическая организация нервных центров выражается в следующем:
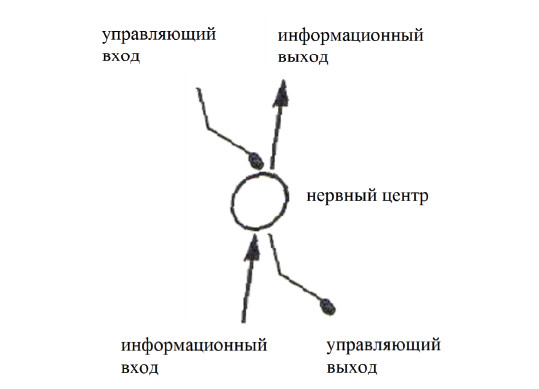
– нижележащий центр является информационным по отношению к вышележащему;
– вышележащий центр является управляющим по отношению к нижележащему.
В результате каждый нервный центр, входящий в определенную иерархическую систему, имеет два входа и два выхода (рис. 16).
Базовые процессы в нервной системе
Нервные центры взаимодействуют посредством двух процессов – возбуждения и торможения. Эти процессы характеризуются определенным набором свойств.
Свойства нервных процессов
1) Возбуждение и торможение могут иррадиировать.
Иррадиация – это распространение процесса из очага возникновения на окружающие клетки. Чем сильнее процесс в очаге возникновения, тем дальше он иррадиирует. Возбуждение иррадиирует быстрее торможения.
2) Иррадиация неизбежно сменяется концентрацией. Концентрация – это сосредоточение процесса в месте его первоначального возникновения. Концентрация происходит медленнее, чем иррадиация.
3) Концентрация сопровождается индукцией. Индукция – это свойство вызывать вокруг себя и после себя противоположный процесс. Индукция вытесняет процесс из той области, на которую он иррадиировал.
4) Возбуждение и торможение могут взаимодействовать. Это происходит при иррадиации процессов. Результат взаимодействия зависит от относительной силы конкурирующих процессов.
Распространение возбуждения и торможения
Возбуждение в нервной системе распространяется самостоятельно, торможение распространяется с помощью возбуждения (рис. 17).
Рис. 17. Схемы, иллюстрирующие процессы распространения возбуждения и торможения в нервной системе
Рис. 18. Варианты возбудительных связей в нервной системе Круг – нейронный модуль, колонка кругов – нервный центр
Тормозные связи могут быть четырех типов. Прямое торможение (рис. 19, А) направлено на уменьшение количества каналов обработки.
Рис. 19. Варианты тормозных связей Стрелками показаны возбудительные связи, треугольник обозначает тормозную связь. Белые кружки – возбудительные нейроны, черные кружки – тормозные нейроны
Реципрокное торможение (рис. 19, Б) обеспечивает согласованную работу систем-антагонистов: активация одной системы сопровождается подавлением другой. Возвратное торможение (рис. 19, В) позволяет ограничить длительность и силу реакции нервной клетки. Латеральное торможение (рис. 19, Г) призвано ограничить распространение возбуждения в нервном центре и подчеркнуть фокус реакции. Эфферентное торможение (рис. 19, Д) позволяет вышележащему центру регулировать активность нейронов нижележащего центра.
Торможение в ЦНС может быть двух видов: постсинаптическое и пресинаптическое (рис. 20). Первое направлено на дендрит или тело клетки, второе – на аксон. Первое изменяет заряд мембраны в сторону гиперполяризации и тем самым снижает возбудимость нейрона. Второе препятствует приходу потенциалов действия к синапсу и тем самым блокирует передачу информации между клетками.
Рис. 20. Виды торможения
Базовые механизмы нервной деятельности
Одной из важных сторон нервной деятельности является формирование адаптивных реакций, вызванных изменениями среды. Эти реакции могут носить врожденный характер и обеспечиваться генетически закрепленными связями. Реакции могут быть и приобретенными. В этом случае они реализуются благодаря образованию новых функциональных связей. Кроме того, адаптивное поведение может носить произвольный характер и выражаться в форме деятельности, направленной на удовлетворение потребностей. В основе реализации адаптивного поведения лежит несколько базовых механизмов нервной деятельности.
Механизм реагирования посредством врожденных связей
Условия обитания закрепили за каждым видом определенный набор врожденных поведенческих реакций. Эти реакции имеют готовую морфофункциональную основу и называются безусловными рефлексами. Безусловный рефлекс – это постоянная, видоспецифическая, стереотипная, генетически закрепленная реакция организма на внутренние или внешние раздражители, осуществляемая при участии нервной системы. Субстратом безусловных рефлексов являются врожденные нервные связи. Рефлекс реализуется путем последовательной активации элементов рефлекторной дуги – афферентного, центрального и эфферентного звена. Запускается рефлекс активацией входа рефлекторной дуги – рецептивного поля данного рефлекса. Безусловные рефлексы обеспечивают выживание вида. Эволюция врожденных форм поведения проявляется в удлинении цепей рефлекторных реакций. Выделяют несколько уровней сложности безусловных рефлекторных реакций: элементарные, координационные, интегративные и сложнейшие.
Элементарные безусловные рефлексы представлены простейшими реакциями, которые вызываются активацией локального поля рецепторов. Они отличаются высокой степенью автоматизма и стереотипности и выражаются в реализации простых движений или в приспособительных изменениях работы отдельных внутренних органов.
Координационные безусловные рефлексы представлены комплексными реакциями, которые запускаются активацией широкого спектра рецепторов. Это синхронные сокращения и расслабления разных мышечных групп, или согласованные изменения работы нескольких внутренних органов. В реализацию таких рефлексов включаются обратные связи. Этот уровень реагирования обеспечивает формирование целостных двигательных актов и гомеостатических реакций.
Интегративные безусловные рефлексы – это комплексные поведенческие акты, которые вызываются биологически значимыми стимулами. Они проявляются в виде сложноорганизованной двигательной активности, которая сопровождается адекватными вегетативными изменениями. На этом уровне реагирования происходит переход от безусловных рефлексов к поведенческим актам.
Единственное, с чем имеет дело мозг, – это информация. Все наши действия руководствуются информацией. Мозг функционирует, пока есть информация. Но что для мозга является информацией? Для мозга информация – это изменение среды, преобразованное в нервный код.
Элементарным обработчиком информации в мозге является нервная клетка, а единицей информации в ЦНС – потенциал действия. Потенциал действия – это электрический импульс, который генерирует нервная клетка и с помощью которого она воздействует на другие клетки. Нейрон кодирует передаваемую им информацию временной последовательностью потенциалов действия (рис. 15).

Рис. 15. Пример записи импульсной активности нервной клетки
Но нейрон – это лишь самый нижний уровень работы с информацией. Нервные клетки в ЦНС объединены в нервные центры. Нервный центр – это скопление нейронов, объединенных общей функцией.
Однако нервные центры – это не монолитные образования. Они подразделяются на модули. Модуль – это функциональное объединение нейронов для совместной обработки информации.
Вместе с тем нервные центры – это не обособленные, не независимые образования. Они объединяются в функциональные системы. Функциональная система – это набор нервных центров, участвующих в выполнении определенной функции.
Таким образом, мы можем выделить несколько уровней функциональной организации ЦНС:
1) нейрон – решающее устройство первого уровня, его задача – кодирование информации;
2) модуль – решающее устройство второго уровня, его задача – обработка информации;
4) функциональная система, ее задача – реализация определенных функций.
Принципы обработки информации
Изучение мозга – это фактически изучение того, как он обрабатывает информацию. Современная наука представляет себе этот процесс в самом общем виде, и выглядит это следующим образом:
– обработка происходит в форме последовательных этапов, каждый этап реализуется определенным нервным центром;
– на каждом из этапов выполняется уникальная операция;
– система функционирует по принципу отрицательной обратной связи: решение нервным центром задачи прерывает его активность.
Такой алгоритм обработки информации возможен благодаря тому, что нервные центры находятся в иерархическом соподчинении. Иерархическая организация нервных центров выражается в следующем:
– нижележащий центр является информационным по отношению к вышележащему;
– вышележащий центр является управляющим по отношению к нижележащему.
В результате каждый нервный центр, входящий в определенную иерархическую систему, имеет два входа и два выхода (рис. 16).
Базовые процессы в нервной системе
Нервные центры взаимодействуют посредством двух процессов – возбуждения и торможения. Эти процессы характеризуются определенным набором свойств.
Свойства нервных процессов
1) Возбуждение и торможение могут иррадиировать.
Иррадиация – это распространение процесса из очага возникновения на окружающие клетки. Чем сильнее процесс в очаге возникновения, тем дальше он иррадиирует. Возбуждение иррадиирует быстрее торможения.
2) Иррадиация неизбежно сменяется концентрацией. Концентрация – это сосредоточение процесса в месте его первоначального возникновения. Концентрация происходит медленнее, чем иррадиация.
3) Концентрация сопровождается индукцией. Индукция – это свойство вызывать вокруг себя и после себя противоположный процесс. Индукция вытесняет процесс из той области, на которую он иррадиировал.
4) Возбуждение и торможение могут взаимодействовать. Это происходит при иррадиации процессов. Результат взаимодействия зависит от относительной силы конкурирующих процессов.
Распространение возбуждения и торможения
Возбуждение в нервной системе распространяется самостоятельно, торможение распространяется с помощью возбуждения (рис. 17).
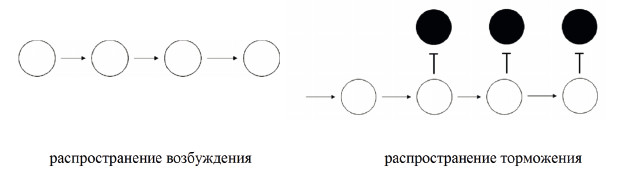
Рис. 17. Схемы, иллюстрирующие процессы распространения возбуждения и торможения в нервной системе
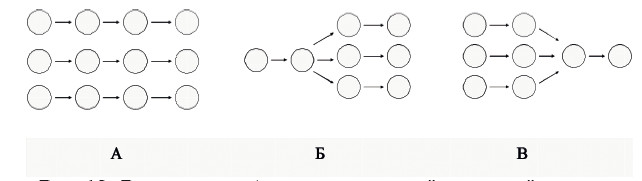
Рис. 18. Варианты возбудительных связей в нервной системе Круг – нейронный модуль, колонка кругов – нервный центр
Тормозные связи могут быть четырех типов. Прямое торможение (рис. 19, А) направлено на уменьшение количества каналов обработки.
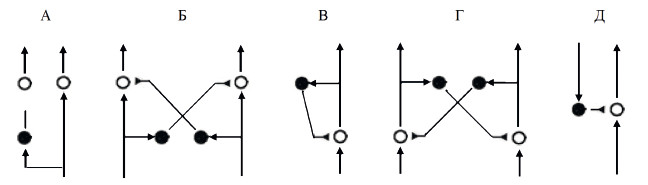
Рис. 19. Варианты тормозных связей Стрелками показаны возбудительные связи, треугольник обозначает тормозную связь. Белые кружки – возбудительные нейроны, черные кружки – тормозные нейроны
Реципрокное торможение (рис. 19, Б) обеспечивает согласованную работу систем-антагонистов: активация одной системы сопровождается подавлением другой. Возвратное торможение (рис. 19, В) позволяет ограничить длительность и силу реакции нервной клетки. Латеральное торможение (рис. 19, Г) призвано ограничить распространение возбуждения в нервном центре и подчеркнуть фокус реакции. Эфферентное торможение (рис. 19, Д) позволяет вышележащему центру регулировать активность нейронов нижележащего центра.
Торможение в ЦНС может быть двух видов: постсинаптическое и пресинаптическое (рис. 20). Первое направлено на дендрит или тело клетки, второе – на аксон. Первое изменяет заряд мембраны в сторону гиперполяризации и тем самым снижает возбудимость нейрона. Второе препятствует приходу потенциалов действия к синапсу и тем самым блокирует передачу информации между клетками.
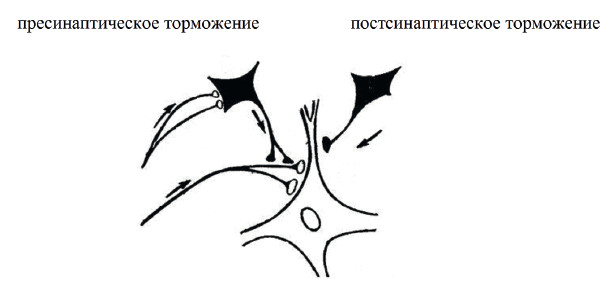
Рис. 20. Виды торможения
Базовые механизмы нервной деятельности
Одной из важных сторон нервной деятельности является формирование адаптивных реакций, вызванных изменениями среды. Эти реакции могут носить врожденный характер и обеспечиваться генетически закрепленными связями. Реакции могут быть и приобретенными. В этом случае они реализуются благодаря образованию новых функциональных связей. Кроме того, адаптивное поведение может носить произвольный характер и выражаться в форме деятельности, направленной на удовлетворение потребностей. В основе реализации адаптивного поведения лежит несколько базовых механизмов нервной деятельности.
Механизм реагирования посредством врожденных связей
Условия обитания закрепили за каждым видом определенный набор врожденных поведенческих реакций. Эти реакции имеют готовую морфофункциональную основу и называются безусловными рефлексами. Безусловный рефлекс – это постоянная, видоспецифическая, стереотипная, генетически закрепленная реакция организма на внутренние или внешние раздражители, осуществляемая при участии нервной системы. Субстратом безусловных рефлексов являются врожденные нервные связи. Рефлекс реализуется путем последовательной активации элементов рефлекторной дуги – афферентного, центрального и эфферентного звена. Запускается рефлекс активацией входа рефлекторной дуги – рецептивного поля данного рефлекса. Безусловные рефлексы обеспечивают выживание вида. Эволюция врожденных форм поведения проявляется в удлинении цепей рефлекторных реакций. Выделяют несколько уровней сложности безусловных рефлекторных реакций: элементарные, координационные, интегративные и сложнейшие.
Элементарные безусловные рефлексы представлены простейшими реакциями, которые вызываются активацией локального поля рецепторов. Они отличаются высокой степенью автоматизма и стереотипности и выражаются в реализации простых движений или в приспособительных изменениях работы отдельных внутренних органов.
Координационные безусловные рефлексы представлены комплексными реакциями, которые запускаются активацией широкого спектра рецепторов. Это синхронные сокращения и расслабления разных мышечных групп, или согласованные изменения работы нескольких внутренних органов. В реализацию таких рефлексов включаются обратные связи. Этот уровень реагирования обеспечивает формирование целостных двигательных актов и гомеостатических реакций.
Интегративные безусловные рефлексы – это комплексные поведенческие акты, которые вызываются биологически значимыми стимулами. Они проявляются в виде сложноорганизованной двигательной активности, которая сопровождается адекватными вегетативными изменениями. На этом уровне реагирования происходит переход от безусловных рефлексов к поведенческим актам.
Сложнейшие безусловные рефлексы (инстинкты) – это комплексы безусловных рефлексов в виде фиксированных последовательностей действий. Инстинкты реализуются благодаря наследственной поведенческой программе, общей для всех представителей вида. Эти реакции отражают исторический опыт вида и запускаются ключевыми стимулами при условии наличия определенной биологической потребности. При реализации инстинкта завершение одной безусловной реакции является сигналом к началу следующей. Этим определяется жесткая последовательность инстинктивных реакций. Достижение данного уровня рефлекторного реагирования обеспечивает переход к сложным формам поведения.
Реагирование посредством врожденных связей – это механизм низшей нервной деятельности. На ранних этапах эволюции нервной системы безусловные реакции – единственная форма реагирования. Круг воспринимаемых раздражителей ограничен, ограничен и репертуар возможных ответных действий. Но безусловные рефлексы не в состоянии в полной мере обеспечить уравновешивание организма со средой. Среда изменчива, а набор врожденных реакций ограничен.
Механизм реагирования посредством приобретенных связей
Реагирование посредством приобретенных связей – это механизм высшей нервной деятельности. Условные рефлексы встраиваются в качестве компонентов в сложные поведенческие акты. Однако условно-рефлекторное обучение является неосознаваемым. Никто из нас не в состоянии сказать, каким набором условных рефлексов он располагает.
Вместе с тем все разнообразие форм поведения невозможно объяснить только лишь рефлекторными реакциями, даже весьма сложными. В ЦНС реализуется еще один механизм, позволяющий сделать поведение бесконечно разнообразным и обеспечивающий наибольший адаптационный потенциал.
Механизм формирования доминанты
Когда достижение результата невозможно путем рефлекторного реагирования, в мозге формируется функциональная система, призванная удовлетворить возникшую потребность. Функциональная система – это временное объединение нервных центров. Оно возможно благодаря образованию доминанты.
Доминанта – это система нервных центров с устойчиво высокой возбудимостью. Это физиологическое проявление мотивации, определяющей направленность поведения. Она создает готовность организма к определенному виду деятельности. Доминанта характеризуется рядом свойств:
– повышенная возбудимость: даже слабые раздражители способны усилить возбуждение доминанты;
– стойкость, инерционность возбуждения: очаг возбуждения сохраняется длительное время, пока не будет удовлетворена потребность;
– способность к суммированию возбуждения: возбуждение центров, не входящих в доминанту, притягивается к доминанте и поддерживает ее возбуждение;
– сопряженное торможение центров-антагонистов: доминанта активно тормозит центры, которые используются для удовлетворения иных потребностей.
Формирование динамичных функциональных систем на основе образования доминант – это физиологический механизм психической деятельности.
Итак, именно благодаря нервной системе организм функционирует как единое целое, решая при этом комплекс важных задач, связанных с удовлетворением биологических, социальных и высших потребностей.
Контрольные вопросы
1. Что в НС является единицей информации?
2. Как по современным представлениям организован в мозге процесс обработки информации?
Начиная от атома и заканчивая Галактикой, в природе нет более сложного объекта, чем человеческий мозг.
Мозг состоит из головного и спинного, и объединяется названием центральная нервная система (ЦНС). ЦНС – это главная управляющая, интегрирующая и информационная система организма, обеспечивающая объединение всех органов в целостный организм, регуляцию всех функций и приспособление организма к окружающей среде.
Различают общие и специфические функции ЦНС.
Общие функции определяются нейронным строением ЦНС, синаптическими связями между нейронами, модульным строением ЦНС. Общие функции базируются на общих механизмах функционирования ЦНС – возбуждении и торможении. Они включают в себя:
1) восприятие, обработку, передачу и хранение информации;
2) преобладание входов над выходами;
3) способность к саморегуляции;
4) параллельную обработку разной информации;
5) функционирование на основе рефлекторно-доминантного принципа.
Специфические функции – это функции, специфичные для каждого отдела мозга (для спинного – свои; для продолговатого – свои и т.д.). Они определяются модулями (ансамблями) , из которых состоят нервные центры. Нервный центр – это совокупность модулей, различных по связям и способам обработки информации, которые регулируют данную функцию и расположены на различных уровнях ЦНС. Модуль – это нейронный комплекс, представляющий собой совокупность локальных нейронных сетей. Размер элементарного модуля по горизонтали составляет 100-150 мкм. Один модуль может входить в состав разных функциональных образований (нервных центров), и сложные паттерны ( pattern , англ. – узор) в нервных центрах определяются скоординированной активностью модулей.
Основные признаки модульной организации ЦНС:
1) локальный синергизм реакции нейронов центральной (ядерной) структуры модуля;
2) наличие тормозной окантовки, образованной клетками с тормозными реакциями на данное раздражение или клетками не реагирующими на него;
3) наличие определённого числа нейронов со стабильными ответами и большого числа нейронов с вариабельными ответами.
Механизм активации модуля . Вначале афферентные импульсы, поступающие в мозг по специфическим и неспецифическим путям, активируют обширную зону мозга, т.е. группу модулей. Затем под влиянием вторичных афферентных сигналов, осуществляющих внутрицентральное взаимодействие, образуется конкретный нейронный модуль, включающий популяцию нейронов с примерным диаметром в 150 мкм. Из таких модулей образуется мозаика нейрональной активности и меняющийся паттерн возбуждения и торможения.
Имеется структурная и функциональная избыточность модульной организации нервных центров. Это связано с возможностью информационной перегрузки нервных центров сенсорными сигналами. Такая избыточность является характерной чертой конструкции нервных центров, что ведёт к их мультифункциональности. Когда индивидуальное участие нейрона в осуществлении какой-либо реакции становится необязательным, а вероятностным, то возможна его заменяемость, что повышает надёжность работы ЦНС. Всё это способствует лёгкости и гибкости перестроек межнейрональных связей, что и объясняет высокую пластичность и большие компенсаторные возможности нервных центров.
Вероятностные модули образуются блоками нейронов, перерабатывающих идентичную информацию. Мозаика этих модулей на всех уровнях ЦНС обеспечивает целенаправленное поведение и образует функциональные системы мозга.
Все методы изучения ЦНС делят на две большие группы:
1) методы экспериментальной нейрофизиологии;
2) методы теоретической нейрофизиологии.

Нейрон является структурной единицей мозга. Функциональной единицей мозга является нейрональный модуль. Число нейронов в мозге человека приближается к 10 12 . Тела нейронов составляют серое вещество мозга. Белое вещество составляют отростки нейронов.
Нейроны – это специализированные клетки, способные принимать, обрабатывать, кодировать, передавать и хранить информацию, организовывать реакцию на раздражения, устанавливать контакты с другими нейронами и клетками.
Классификация нейронов.
Нейроны классифицируются по различным признакам.
1. По размерам и форме различают овальные, звёздчатые, корзинчатые, пирамидные, многоугольные, грушевидные, нейроны-зёрна и др. Размеры тел нейронов колеблются от 5 до 200 мкм.
2. По отросткам нейроны делятся на:
1) униполярные;
2) биполярные;
3) ложноуниполярные;
4) мультиполярные.
3. По функции нейроны делятся на:
1) афферентные (осуществляют восприятие и проведение сигнала, идущего с рецептора);
2) вставочные (выполняют различные функции, в том числе осуществляют передачу сигнала от одного нейрона к другому; способствуют распределению сигналов по нейронным сетям; осуществляют торможение (тормозные нейроны); постоянно поддерживают активность отдельных нервных центров (пейсмекерные нейроны); осуществляют принятие решения (командные нейроны));
3) эфферентные:
- мотонейроны;
- нейроны автономной нервной системы.
4. По нейрональной активности нейроны можно разделить на:
1) нейроны с фоновой (автоматической) активностью, которая может быть единичной (в редких случаях); пачковой (2-20 Гц) и групповой (более 20 Гц);
2) нейроны с вызванной активностью:
- on-нейроны (генерируют импульсы при включении раздражителя);
- off-нейроны (генерируют импульсы при выключении раздражителя);
- on-off-нейроны (генерируют импульсы при включении и выключении раздражителя).
5. По характеру воспринимаемых раздражений нейроны делятся на:
1) моносенсорные;
2) бисенсорные;
3) полисенсорные;
4) нейроны новизны (возбуждаются только в ответ на незнакомые раздражения).
Функции нейрона:
1) возбудимость;
2) проводимость;
3) переработка информации;
4) интеграция информации.
Для осуществления этих функций нейрон имеет:
1) воспринимающую часть – это дендриты и мембрана сомы;
2) интегративную часть – это сома с аксонным холмиком;
3) передающую часть – это аксонный холмик с аксоном.
Строение нейронов.
Нейрон состоит из тела и отростков (дендриты и аксон).
Тело нейрона (сома) выполняет интегративную функцию и трофическую относительно своих отростков и их синапсов функции. Сома обеспечивает рост дендритов и аксона. Она заключена в многослойную мембрану, обеспечивающую формирование и распространение электротонического потенциала к начальному сегменту , который вместе с аксоном обеспечивают передающую функцию. Характерной особенностью мембраны этого участка является высокая плотность на ней натриевых потенциалзависимых каналов.
Дендриты – это отростки нейрона, выполняющие воспринимающую функцию. Их мембрана способна реагировать на медиаторы, выделяемые аксонными окончаниями, изменением электрического потенциала. Обычно нейрон имеет несколько ветвящихся дендритов, что обусловлено необходимостью адекватного потока информации. Она поступает через специализированные контакты, так называемые шипики. За счёт шипиков значительно возрастает воспринимающая поверхность нейронов, а соответственно увеличивается поток информации в нейрон. Чем сложнее функция нервной системы, тем больше шипиков на дендритах нейронов. Если какая-либо группа шипиков перестаёт получать информацию, то они исчезают.
Аксон представляет собой вырост цитоплазмы, приспособленный для проведения и передачи информации. Аксон начинается от аксонного холмика, имеет постоянный диаметр, в большинстве случаев одет в миелиновую оболочку, образованную глией. Передающую функцию аксон осуществляет через разветвлённые окончания, в которых находятся митохондрии и секреторные образования.
На одном нейроне может быть до 10 000 синапсов, через которые он получает возбуждение и тормозные воздействия от других нейронов. Нейрон может находиться в различных состояниях:
1) в состоянии покоя , когда практически отсутствуют колебания мембранного потенциала, потенциал действия не генерируется;
2) в состоянии активности , когда нейрон генерирует потенциалы действия.
Состояние активности может быть индуцировано за счёт поступления к нейрону импульсов от других нейронов или быть спонтанным (автоматия). В этом случае нейрон играет роль пейсмекера ( pace , англ. – шаг; maker , англ. – измеритель) – водителя ритма. Такие нейроны имеются в ряде центров, например, в центре дыхания. В состоянии торможения оно проявляется в том, что нейрон, прекращает свою импульсную активность (нейрон-пейсмекер, или нейрон, получающий возбуждающие воздействия). В основе торможения лежит явление гиперполяризации нейрона (это характерно для постсинаптического торможения).
Обычно нейрон посылает потенциал действия по аксону к другому нейрону или к эффектору, например, к мышце. Такой путь проведения называется ортодромным ( orthos , гр. – прямой; dromos , гр. – путь). Если сигнал распространяется на сому и дендриты, то это антидромное ( anti , гр. – противо-; dromos , гр. – путь) распространение возбуждения.

Интегративная функция нейрона.
Под интегративной функцией нейрона подразумевается не только количественная трансформация поступающих сигналов, но и их качественные преобразования. Существуют две теории, объясняющие эту функцию.
Мембранная теория (теория электрической суммации) (Экклс, Катц, Стивенсон, 1950). Согласно этой теории мембрана нейрона представляется сумматором ВПСП и ТПСП. При электрической суммации приходящих сигналов она интегрирует входящую информацию, генерируя ПД, который затем поступает с аксонного холмика по аксону на периферию.
В последнее время оказалось, что между функцией, структурой и метаболизмом нейрона имеется связь (П.К.Анохин). При возбуждении нейрона регистрируются изменения в ядерном комплексе, цитоплазме и нейротрубочках. Обнаружены также изменения обмена белков, РНК и углеводов в нейроплазме. Эти данные позволили академику П.К.Анохину в 1970 г выдвинуть химическую теорию интегративной функции нейрона. Согласно этой теории интеграция информации происходит не только с помощью электрических процессов на мембране нейрона, но и с помощью нейрохимических преобразований. Косвенным доказательством справедливости химической теории является сложность строения нейрона.
Гематоэнцефалический барьер. Для мозга очень характерно наличие специфического барьера для многих веществ, находящихся в крови, в том числе для гормонов, биологически активных веществ, некоторых антибиотиков и др. Это, с одной стороны, обусловлено наличием особого устройства капилляров мозга: они мало проницаемы для многих веществ вследствие того, что их эндотелий является эндотелием плотного типа, в таком эндотелии мало пор, контакты между соседними эндотелиальными клетками плотные и не пропускают молекулы через этот барьер. Кроме этого отростки глиоцитов образуют на поверхности капилляров слой, отграничивающий нейроны от непосредственного соприкосновения с сосудистой стенкой.
Всё это обеспечивает стабильность работы нейрона. Гематоэнцефалический барьер сохраняет своё значение (и свои механизмы) в тех местах, где образуется ликвор, т.е. в сосудистых образованиях желудочков мозга. Ликвор – это жидкость с минимальным содержанием в ней молекул биологически активных веществ и гормонов. Ликвор необходим для обеспечения гидравлической подушки мягким тканям мозга. Одновременно ликвор осуществляет функции лимфы, с помощью ликвора осуществляется дренирование тканей мозга и удаление из них осколков клеток, больших молекул и т.п. в венозную систему, куда впадает ликвор.
Рефлекс и рефлекторная дуга.
Рефлекс – это универсальная форма взаимодействия организма со средой в виде ответной реакции, возникающей на раздражение рецепторов и осуществляемой с обязательным участием ЦНС.
Рефлекторная дуга – это морфологическая основа рефлекса, включающая следующие звенья:
1) рецептивное поле рефлекса – это определённый участок воспринимающей чувствительной поверхности организма с расположенными здесь рецепторными клетками, которые трансформируют энергию раздражения в энергию нервного импульса;
2) афферентное звено – это афферентные нервы, назначение которых состоит в проведении нервных импульсов в ЦНС;
3) нервный центр (центральное звено) – это совокупность центральных нервных структур, отвечающих за приём, обработку (анализ, синтез, интеграция и др.) и передачу нервных импульсов на эфферентное звено. В простейшем случае центральное звено имеет один синапс, где нервные импульсы переключаются с афферентного пути на эфферентный нейрон – это моносинаптическая рефлекторная дуга (классический пример – спинальные миотатические (возникающие в ответ на растяжение мышцы) рефлексы). Если в центральном звене имеется один или несколько вставочных нейронов, то такая дуга называется полисинаптической;
4) эфферентное звено – это эфферентные нервы, по которым нервные импульсы передаются на эффектор (рабочий орган);
5) эффектор .
Классическая схема рефлекторной реакции – это простая прямая связь: стимул → нервный центр → ответная реакция. Современные представления о рефлексе как о целесообразной реакции организма диктуют необходимость дополнить рефлекторную дугу ещё одним звеном – обратной связью (шестое звено рефлекторной дуги). Она устанавливает связь между полученным результатом рефлекса и нервным центром, который выдал команду на достижение этого результата. Обратная связь принципиально реализуется многими принципиально одинаковыми механизмами, суть которых состоит в передаче информации от рабочего органа к нервному центру. В результате этого рефлекторная дуга превращается в рефлекторное кольцо . Так формируется самонастраивающийся нервный контур регуляции физиологических функций – основа функциональной системы (П.К.Анохин).
Время рефлекса (латентный период рефлекса) – это время от момента нанесения раздражения до конечного эффекта. В моносинаптическом рефлексе оно достигает 20-25 мс. Это время расходуется на возбуждение рецепторов, проведение возбуждения по афферентным волокнам, передача возбуждения с афферентных нейронов на эфферентные (возможно через несколько вставочных), проведение возбуждения по эфферентным волокнам и передача возбуждения с эфферентного нерва на эффектор. Центральное время рефлекса – это промежуток времени, за который нервный импульс проводится по структурам мозга. В случае моносинаптической рефлекторной дуги оно составляет примерно 1,5-2 мс – это время, необходимое для передачи возбуждения в одном синапсе. Таким образом, центральное время рефлекса косвенно указывает на число синаптических передач, имеющих место в данном рефлексе. Центральное время у полисинаптических рефлексов более 3 мс. В целом, полисинаптические рефлексы очень широко распространены в организме человека. Центральное время рефлекса является главной составляющей общего времени рефлекса.
Классификация рефлексов.
Единой классификации рефлексов нет. Отметим наиболее существенные подходы к классификации рефлексов.
1. По способу образования рефлекторной дуги (И.П.Павлов) рефлексы делятся на безусловные и условные. В случае безусловных рефлексов рефлекторная дуга формируется независимо от жизненного опыта индивидуума, т.е. становление безусловных рефлекторных дуг генетически запрограммировано. Условно-рефлекторный процесс требует создания новых рефлекторных дуг на основе сочетания работы безусловных рефлекторных дуг и индифферентного раздражителя.
2. По биологическому значению приспособительного результата выделяют питьевые, пищевые, половые, оборонительные, ориентировочные, локомоторные рефлексы, рефлексы положения и др.
3. В зависимости от вида рецепторов, с которых начинаются рефлексы, они подразделяются на:
1) экстерорецептивные ;
2) интерорецептивные ;
3) проприоцептивные .
4. По расположению нервных центров, в которых замыкаются рефлексы, они подразделяются на:
1) спинальные ;
2) бульбарные ;
3) мезенцефалические ;
4) диэнцефалические ;
5) кортикальные .
5. По типу эффектора рефлексы можно разделить на:
1) двигательные;
2) сосудодвигательные;
3) секреторные.
6. Рефлексы соматической и автономной нервной системы соответственно подразделяются на соматические и вегетативные.
7. С учётом уровня интегративной деятельности мозга А.Б.Коган различает следующие виды рефлексов:
1) элементарные безусловные рефлексы – это простые рефлекторные реакции, осуществляемые на уровне отдельных сегментов спинного мозга по жёстко детерминированным программам с высокой степенью автоматизма и стереотипности;
2) координационные безусловные рефлексы – это согласованные друг с другом акты локомоторной деятельности, или комплексные реакции вегетативных функциональных объединений внутренних органов;
3) интегративные безусловные рефлексы – это комплексные поведенческие акты в тесной связи с вегетативным обеспечением системного характера, имеющие определённое биологическое значение;
4) сложнейшие безусловные рефлексы (инстинкты) – это видовые стереотипы поведения, реализующиеся на базе интегративных рефлексов по генетически заданной программе;
5) элементарные условные рефлексы – это интегративные реакции опережающего характера, вызываемые раздражителями, имеющими сигнальное значение, приобретённое в результате жизненного опыта, что делает их биологически значимыми;
6) сложные формы высшей нервной деятельности – это психические реакции, возникающие на основе интеграции элементарных условных рефлексов и аналитико-синтетических механизмов абстрагирования. Однако следует отметить, что далеко не все психические реакции возможно объяснить рефлекторной деятельностью.

Потоки импульсов – сигналы о состоянии внешней и внутренней среды – вызывают различные рефлексы, обеспечивающие текущую деятельность организма и его приспособление к меняющимся условиям среды. В каждый момент времени необходимы не все рефлексы, а определённые, которые уравновешивают организм со средой, и это достигается координацией, т.е. согласованностью рефлексов, т.е. торможением одних и преобладанием других.
Координация (co, лат. – вместе + ordinatio , лат. – расположение в порядке = согласование) рефлексов – это взаимодействие нервных процессов в ЦНС, обеспечивающее её согласованную деятельность. Координация способствует реализации всех функций ЦНС и проявляется различными эффектами – это может быть облегчение или угнетение возбуждения. Реализация координационной деятельности происходит на основе следующих принципов.
Центральная нервная система координирует деятельность всех органов и систем, обеспечивает эффективное приспособление организма к изменениям окружающей среды, формирует целенаправленное поведение. Эти жизненно важные задачи решаются благодаря интегративной деятельности ЦНС.
Интегративная деятельность ЦНС – это ее способность объединять, обобщать все поступающие сигналы, отрабатывать их в связи с прошлым опытом. В результате формируется определенная реакция организма биологически или социально наиболее важная в данной ситуации.
В интегративной деятельности условно выделяют 4 основных уровня:
1. Интеграция на уровне рецептора. Рецептор, воспринимая информацию, осуществляет ее первичный отбор по интенсивности, продолжительности, модальности и формирует нервные импульсы.
2. Интегративная деятельность нейрона – это способность нейрона воспринимать возбуждение и торможение, обрабатывать их с учетом генетической и приобретенной памяти нейрона и вырабатывать временную последовательность потенциалов действия. Интегративная деятельность нейрона базируется на конвергентных свойствах нейрона и его структурных изменениях, лежащих в основе обучения и памяти.
3. Интеграция на уровне центра. Нервный центр – это совокупность нервных клеток, расположенных на различных уровнях ЦНС и обеспечивающих определенную физиологическую реакцию организма. Иерархический принцип строения нервных центров создает возможность тонко дифференцировать ответные реакции.
Свойства нервных центров обусловлены свойствами нейронов, центральных синапсов и типами связей между нейронами. Для нервных центров характерны:
· высокий аэробный обмен веществ и высокая чувствительность к гипоксии;
· чувствительность к фармакологическим препаратам;
· меньшая возбудимость, чем у нервных волокон;
· односторонняя передача возбуждения;
· последействие (продолжение рефлекса после прекращения действия раздражителя);
· суммация (способность центральных синапсов к суммации допороговых импульсов и явлениями облегчения и конвергенции).
4. Межцентральная интеграция обеспечивает согласование деятельности различных нервных центров и формирование сложных поведенческих, эмоциональных и адаптивных реакций, организовывая деятельность организма как единого целого.
В естественных условиях любой рефлекторный акт является результатом интегративной деятельности. В основе интегративной деятельности ЦНС лежат механизмы координации.
Координация – это согласованное взаимодействие процессов возбуждения и торможения в ЦНС. Координация процессов в нервных центрах происходит при осуществлении любого рефлекторного акта. Этот процесс базируется на принципах конвергенции, дивергенции и обратной связи (рисунок 13).
Конвергенция – схождение различных путей (тормозящих, возбуждающих) проведения нервных импульсов на одной нервной клетке. Это обуславливает интегративную функцию нейрона. Принцип конвергенции лежит в основе таких процессов, как общий конечный путь, пространственная суммация и окклюзия (см. ниже).
Дивергенция – это способность нейрона устанавливать многочисленные синаптические связи с другими нервными клетками. Благодаря процессу дивергенции один нейрон может участвовать в различных нервных реакциях и контролировать большое число других нейронов, а также каждый нейрон может обеспечивать широкое перераспределение импульсов, что приводит к иррадиации возбуждения.
Рис. 13. Схема дивергенции (А) и конвергенции (Б) сигналов в ЦНС. Схематически изображены нервные клетки, их аксоны и образуемые ими синапсы. Стрелки отмечают направление передачи сенсорной информации
Обратные связи – поступление нервных импульсов в ЦНС с иннервируемого органа или клетки. Обратные связи разделяют на центральные (кольцевой тип связи между нейронами) и рефлекторные (импульсы возвращаются в нервный центр с рецепторов иннервируемого органа). По эффекту обратные связи могут быть положительными и отрицательными.
В нервные центры от рецепторов обычно поступает ритмическая импульсация. При этом ответная реакция ЦНС не всегда линейно зависит от силы и частоты раздражителя. В нервных центрах можно наблюдать явление суммации допороговых стимулов и окклюзии сверхпороговых.
Суммация. Различают пространственную и последовательную суммацию. Последовательная суммация возникает при ритмической стимуляции одного рецептивного поля. В основе ее лежит механизм облегчения. Пространственная суммация допороговых стимулов происходит при одновременной стимуляции различных рецептивных полей. Она базируется на принципах облегчения и конвергенции.
Окклюзия- это процесс, при котором общая ответная реакция нервных центров на сверхпороговые стимулы меньше, чем алгебраическая сумма раздельных эффекторов каждого. Последовательная окклюзия происходит при быстрой повторной стимуляции нейрона. При этом нейрон не воспроизводит все поступающие к нему сигналы, в результате чего происходит уменьшение сильных сигналов.
В основе пространственной окклюзии лежит процесс конвергенции, который приводит к уменьшению количества суммарнореагирующих нейронов.
Различные рефлекторные реакции могут взаимодействовать между собой. Примером такого взаимодействия являются феномены иррадиации возбуждения, доминанта и принцип общего конечного пути.
Общий конечный путь. Данный принцип введен в физиологию Ч. Шеррингтоном и основан на способности различных проводящих путей создавать синаптические контакты на одной и той же эффекторной клетке. В основе этого лежит принцип конвергенции. К мотонейронам спинного мозга кроме первичных афферентных волокон конвергируют волокна различных нисходящих трактов, идущих из центральных структур мозга, а также аксоны возбуждающих и тормозных вставочных нейронов спинного мозга. Вследствие этого Ч. Шеррингтон именно мотонейроны рассматривал как общий конечный путь многочисленных структур мозга, связанных с регуляцией моторных функций. Принцип общего конечного пути показывает, каким образом одна и та же конечная реакция может быть получена при раздражении различных структур мозга. Этот принцип имеет важное значение для анализа рефлекторной деятельности нервной системы.
Иррадиация возбуждения – это широкое распространение возбуждения по различным нервным центрам. В основе этого процесса лежит большая продолжительность и сила поступающих стимулов в ЦНС, высокая возбудимость нейронов и ослабление центрального торможения. Механизм иррадиации – дивергенция.
Иррадиация возбуждения по нервным центрам способствует возможности одних нейронов участвовать в различных нервных реакциях и контролировать деятельность других нейронов. Однако иррадиация возбуждения может стать патологической в связи с возникновением сильного очага возбуждения и с изменением свойств нервной ткани, усиливающим распространение возбуждения по ней, как это бывает при эпилепсии.
Доминанта – временно господствующий очаг возбуждения в ЦНС, обуславливающий интегральный характер функционирования нервных центров в каждый данный период времени и определяющий целесообразное поведение человека.
Доминантный очаг возбуждения притягивает к себе возбуждение из других нервных центров и одновременно подавляет их деятельность. Доминантный очаг обладает также и свойством притягивания сигналов с других рецептивных полей.
Доминанта может исчезнуть при возникновении более сильной доминанты, реализации доминантного рефлекса или ее затухания в следствие истощения энергетических ресурсов.
Различают следующие виды доминанты: физиологическую и патологическую. Физиологическая доминанта обусловлена биологическими и социальными потребностями (мотивами). Патологическая – проявляется в виде вредных привычек (табакокурение, алкоголизм, наркомания) или при психических расстройствах.
И. П. Павлов указывал также на то, что принцип доминанты лежит в основе формирования временной функциональной связи (условного рефлекса). Таким образом, явление доминанты является одним из важнейших принципов работы ЦНС.
Тема 3. Частная физиология центральной нервной системы.
Вопросы для самоподготовки.
2. Функции спинного мозга.
2.1. Проводниковая функция (проводящие пути спинного мозга).
2.2. Рефлекторная функция. Рефлексы спинного мозга.
2.3. Спинальный шок.
II. Продолговатый мозг.
1. Строение продолговатого мозга (границы, пирамиды, оливы, особенности расположения белого и серого вещества, ядра продолговатого мозга).
2. Функции продолговатого мозга.
2.1. Проводниковая функция продолговатого мозга.
2.2. Рефлекторная деятельность продолговатого мозга:
- центры жизненноважных рефлексов (дыхательный, сердечно-сосудистый);
- защитные рефлексы (мигание, чихание, кашель, рвотный акт и др.);
- рефлексы, связанные с пищеварительной деятельностью (глотание, отделение пищеварительных соков и др.);
- рефлексы, связанные с функциями ядер черепно-мозговых нервов, расположенных в продолговатом мозге (в том числе и вегетативные);
3. Участие продолговатого мозга в регуляции мышечного тонуса и рефлексов позы.
III. Средний мозг.
1. Строение среднего мозга (границы, ядра).
2. Функции среднего мозга.
2.1. Проводниковая функция.
2.2. Рефлекторная деятельность среднего мозга:
- роль красных ядер и черной субстанции в распределении мышечного тонуса (синдром Паркинсона и децеребрационная ригидность).
3. Роль среднего мозга в сохранении нормального положения тела в пространстве (выпрямительные и статокинетические рефлексы).
IV. Ретикулярная формация ствола мозга.
- строение РФ (расположение, ядра РФ, афферентные и эфферентные связи, виды нейрональных контактов);
- функции РФ (контроль сна и бодрствования, участие в регуляции вегетативных функций, фазный и тонический мышечный контроль, участие в механизмах формирования целостных условно -рефлекторных реакций организма).
1. Строение мозжечка (червь, полушария, кора и белое вещество, ножки, ядра; связи мозжечка с другими структурами ЦНС).
2. Функции мозжечка:
- участие в координации движений;
- регуляция мышечного тонуса;
- сохранение позы и равновесия тела;
- участие в регуляции вегетативных функций (функций внутренних органов);
- влияние мозжечка на образование условных рефлексов.
3. Симптомы нарушения функций мозжечка (астазия, атаксия, астения, атония, дистония и др.).
VI. Промежуточный мозг.
1. Составные части промежуточного мозга (эпиталамус, метаталамус, таламус и гипоталамус).
2.1. Нейронная организация.
2.1. Функции таламуса:
- роль специфических (переключательных и ассоциативных) ядер, моторных и неспецифических ядер;
- регуляция важных физиологических состояний (смена сна и бодрствования, сохранение сознания, развитие процессов внутреннего торможения и др.).
3.1. Нейронная организация.
3.2. Афферентные и эфферентные связи гипоталамуса.
3.3. Функции гипоталамуса:
- роль гипоталамуса в регуляции вегетативных функций;
- участие в регуляции поведенческих реакций;
3.4. Функциональные расстройства у людей с повреждениями гипоталамуса.
VII. Базальные ядра.
1. Структуры, входящие в состав базальных ядер и их связи.
2. Функции базальных ядер:
- обеспечение миостатических реакций (плавности движений);
- обеспечение автоматизма движений – бессознательного их выполнения;
- обеспечение движения мимических мышц и участие в формировании эмоциональных реакций;
- формирование защитных ориентировочных рефлексов.
3. Патофизиология базальных ганглиев:
- гипофункция медиаторных систем (болезнь Паркинсона);
- гиперфункциональные симптомы (ригидность, баллизм, атетоз, хорея, тремор).
VIII. Кора больших полушарий.
1. Организация коры больших полушарий (клеточные слои, доли, области, поля).
2. Древняя и старая кора.
2.1. Структуры, входящие в состав (обонятельный мозг и лимбическая область).
2.2. Функции древней и старой коры:
- обеспечение реакций настораживания и внимания;
- регуляция вегетативных функций;
- осуществление видоспецифического (инстинктивного) поведения;
- осуществление социального поведения;
- участие в процессах сохранения памяти.
3. Функции новой коры.
3.1. Чувствительные зоны коры большого полушария.
3.2. Моторные зоны коры большого полушария.
3.3. Электрические явления в коре больших полушарий (электроэнцефалография – ЭЭГ).
Читайте также: