
Как называется процесс согласования активности мышц тела
Обновлено: 16.05.2024
Большое значение в клинике имеет процесс реабилитации пациентов после травм различного характера. Для более правильного подхода к назначению упражнений для адаптации мышц к нагрузке необходимо учитывать различные аспекты данного процесса: варианты его протекания, генетические и морфофизиологические возможности пациента.
Адаптация мышечного волокна — это приспособление волокон мышц к нагрузке, которое лежит как на фенотипическом, так и на генотипическом уровнях. Скелетные мышцы неоднородны по своему составу (табл. 1), следовательно, для достижения полноценного развития необходимо сочетать аэробные и анаэробные нагрузки, так как различные волокна отвечают на них по-разному [1—6] (табл. 2).
Таблица 1. Основные типы мышечных волокон [5—9]. Таблица 2. Различие между аэробными и анаэробными упражнениями [9—12].
Примечание. Аэробные нагрузки в основном являются статистическими (мышцы находятся в постоянном напряжении), анаэробные — динамическими (происходит чередование напряжения и расслабления мышц). Полный расход нейромедиатора в синапсах характерен для продолжительных динамических нагрузок, а наиболее низкий расход — для статических нагрузок [13, 14]. Кроме того, в волокнах I типа более выражены эндотелийзависимая дилатация и чувствительность к катехоламинам [15—19].
Пути адаптации мышечного волокна к нагрузке можно разделить на два типа: компенсаторный и биохимический.
Компенсаторная адаптация: может достигаться за счет увеличения количества саркоплазмы (именно она передает напряжение с волокон на сухожилия [20]) или за счет увеличения количества миофибрилл [21] и позволяет выполнять бо́льшую физическую работу [20—23].
Саркоплазматическая адаптация. Для развития волокон по данному типу необходимо применение аэробных нагрузок, которые ведут к изменению количества и качества митохондрий в мышце: кристы уплотняются, а также не только увеличиваются в количестве и размере (хотя наблюдается деградация некоторых), но и объединяются в цепочки [24—27]. Кроме того, при аэробных нагрузках и потреблении кислорода на 80% от максимально возможного наблюдается прирост митохондрий в волокнах I типа (с исходно высоким окислительным потенциалом), повышение нагрузки не приводит к изменениям. Потребление кислорода на 80—95% от максимально возможного характеризуется приростом окислительного потенциала в волокнах II типа (с исходно низким окислительным потенциалом), при меньших нагрузках изменений не наблюдалось [1, 19]. Так, по завершению длительного курса тренировок было выявлено увеличение количества волокон IIA типа. Увеличение количества митохондрий в свою очередь инициирует рост миофибрилл [28—31]. В результате растет способность мышцы выдерживать статическую или высокоинтенсивную нагрузку. Происходит миофибриллярная адаптация — анаэробная адаптация к силовой нагрузке. Увеличение мышечных волокон происходит за счет увеличения количества миофибрилл [32].
Чрезвычайно важны также генотипические изменения при адаптации [33, 34]. При нагрузках повышается экспрессия PI3K, который активирует экспрессию гена Akt — ключевого в гипертрофии (при нагрузках) и атрофии (падение экспрессии в отсутствие нагрузок [35]). В дальнейшем активируется фермент mTOR [36] (в мышцах содержится в виде двух комплексов — mTORC1 и mTORC2 [37—40]), регулятором которого служит фосфатидная кислота, вырабатывающаяся в мышцах при их работе [39]. Суть действия фермента — повышение отношения синтез/распад белка, что приводит к гипертрофии. Так, путь PI3K—Akt—mTOR является ключевым в гипертрофии мышечного волокна, следовательно, при его блокаде (рапамицин) данный процесс прекращается [41—43]. Отрицательным же регулятором данного процесса служит AMPK [44, 45].
Для полноценного функционирования мышц им необходимо не только более чем 80% потребление кислорода, но и достаточное количество питательных веществ, необходимых для синтеза новых мышечных белков и волокон, в частности, мясо, яйца, жирные кислоты — Омега-3, специальные смеси аминокислот [46—48].
Второй тип адаптации — биохимический. Он обеспечивается более полным использованием субстрата, увеличивая емкость реакций энергообеспечения [36, 48, 49].
Показателем развития мышечного волокна служит количество фермента креатинфосфокиназы (КФК) [50, 51]. Ее активность у нетренированных людей снижена. У них также не выявлено изменений концентрации КФК в процессе нагрузки, в отличие от спортсменов (наблюдается резкий скачок) [51—56]. Активность КФК свидетельствует о том, что у спортсменов (в отличие от неспортивных людей) наряду с активацией гликолиза задействован и креатинфосфокиназный механизм энергообразования, установлено повышение емкости креатинфосфатного механизма образования энергии в мышечной ткани [51, 57]. Этим объясняется скачок активности КФК после физической нагрузки [51, 58].
Еще один важный показатель — концентрация лактата. Он служит показателем анаэробных процессов энергообразования при мышечной работе. У людей, не занимающихся спортом, наблюдается повышение уровня молочной кислоты [51, 59] до и после нагрузки примерно в 8,68 раза, тогда как у спортсменов происходит увеличение примерно в 2,38 раза. Это объясняется более экономным режимом работы скелетной мускулатуры второй группы, обусловленным тренировками. Активность лактатдегидрогеназы (ЛДГ), в свою очередь, до нагрузки у спортсменов снижена, что указывает на меньшую повреждаемость мышцы действием фоновой физической нагрузки (ходьба). После нагрузки было отмечено повышение активности фермента в крови: у спортсменов в 1,8 раза, у нетренированных людей — в 1,2 раза [51]. Примечательно, что концентрация ЛДГ в крови у тренированных людей может повышаться в 2—10 раз вследствие разрушения клеточных мембран и высвобождения ЛДГ из клеток [28, 41—43, 48].
Развитие этих систем необходимо для нивелирования усиления свободнорадикальных реакций, необходимых для перестройки энергетического обмена [12, 60—64] на уровне организма в условиях плановых физических нагрузок. В условиях умеренных физических нагрузок из-за повышенных энергетических затрат увеличивается потребление кислорода и наступает состояние физиологической гипоксии [63—66], следовательно, образуются реактивно-активные формы кислорода с последующим включением свободнорадикальных и перекисных реакций путем мобилизации эндогенных жиров и стимуляции симпатико-адреналовой системы [49, 67—71]. В свою очередь образовавшийся эндогенный кислород обеспечивает поддержание интенсивного энергетического обмена [36, 39, 40, 71—76].
Заключение
Необходимо отметить чрезвычайную важность понимания способов и путей адаптации мышечного волокна к нагрузке для процесса регенерации после травм. Несмотря на малую изученность генотипической адаптации мышечных волокон к нагрузке уже сейчас необходимо составлять реабилитационные программы на их основе; тренировать спортсменов и вести их отбор, базируясь на основах данных процессов. Необходимо использовать эти знания для коррекции как программ реабилитации пациента и индивидуальных программ тренировок, так и для усовершенствования спорта в целом.
Даны определения адаптации, стресса (общего адаптационного синдрома) и суперкомпенсации. Рассмотрены виды адаптации (срочная и долговременная) и условия адаптации скелетных мышц человека к физическим нагрузкам.
Адаптация скелетных мышц человека к физическим нагрузкам
Организм человека устроен таким образом, что, попадая в новые для него условия, он может к ним приспособиться. Такое свойство организма человека получило название адаптация.
Понятия адаптации и стресса
Стресс – неспецифическая (общая) реакция организма на воздействие, нарушающее его гомеостаз.
Г. Селье установил, что на разные по качеству, но сильные раздражители (стресс) организм для выравнивания гомеостаза всегда отвечает однотипными реакциями, которые были названы им общим адаптационным синдромом.
Спортивную тренировку можно рассматривать как адаптацию организма спортсмена к нагрузкам определенной направленности. При этом под воздействием систематических тренировочных нагрузок внутренняя среда организма человека претерпевает значительные изменения. После прекращения нагрузки в организме начинаются процессы, направленные на восстановление исходного состояния.
Виды адаптации
Различают срочную и долговременную адаптацию организма спортсмена к тренировочным воздействиям. Так как основным объектом этой статьи являются скелетные мышцы, вопросы адаптации будут рассматриваться в этом ракурсе.
Срочная адаптация – это структурно-функциональная перестройка, происходящая в организме спортсмена непосредственно во время выполнения физических упражнений. Основной целью срочной адаптации является создание оптимальных условий для функционирования мышц, прежде всего, за счет увеличения их энергоснабжения. В связи с этим, значительно ускоряются реакции катаболизма[2] при одновременном снижении скорости анаболических[3] процессов (в основном, синтеза белков), так как в ходе катаболических реакций выделяется энергия.
Долговременная адаптация – структурно-функциональная перестройка, происходящая в организме в ответ на длительное или многократное воздействие физической нагрузки. Долговременная адаптация протекает в организме спортсмена в промежутках между тренировками.
Условия адаптации
Первым условием является многократное (повторное) применение физических нагрузок. Однократная физическая нагрузка не вызывает стойких адаптационных перестроек в организме. Если же физические нагрузки повторяются, в организме создается необходимый метаболический фон, который обеспечивает постепенность формирования морфологических, биохимических и функциональных изменений. При повторяющихся тренировочных нагрузках благодаря активации генетического аппарата мышечных волокон в мышцах увеличивается содержание структурных и сократительных белков, вследствие чего мышцы становятся более резистентными к задаваемой нагрузке.
Вторым условием, определяющим процесс адаптации организма к физическим нагрузкам, является их регулярное применение Необходимость регулярно выполнять физические упражнения связана с изменениями метаболизма, которые происходят в организме в процессе физических нагрузок. В зависимости от интенсивности и длительности физической нагрузки, в организме происходят изменения в обмене веществ, которые могут быть ограничены локальными сдвигами в энергетическом обмене или затрагивать метаболизм всего организма. В последнем случае процесс восстановления метаболизма до уровня покоя занимает значительно больше времени и требует большего периода отдыха.
Долговременная адаптация организма к различным факторам внешней среды возможна благодаря явлению суперкомпенсации (сверхвосстановления). В области мышечной деятельности это явление первыми описали Л.И. Ямпольский (1949) и Н.Н. Яковлев (1949,1955). В основе суперкомпенсации лежит взаимодействие между нагрузкой и восстановлением. Это – циклический процесс и его пусковым стимулом является физическая нагрузка. После нагрузки возникает утомление и резкое снижение работоспособности спортсмена, что соответствует первой фазе цикла. Во второй фазе начинается процесс восстановления работоспособности. К концу второй фазы работоспособность спортсменов достигает исходного уровня. В третьей фазе (фазе суперкомпенсации) работоспособность превышает исходный уровень. Последняя, четвертая фаза характеризуется возвращением работоспособности к исходному уровню (рис.1 а).
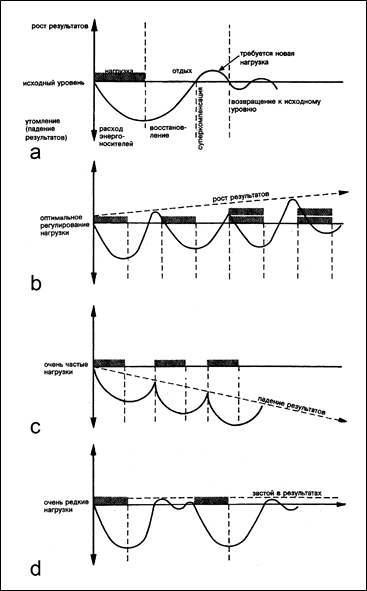
Очень наглядно эффект суперкомпенсации виден на примере восстановления энергетических запасов мышц. Перед началом тренировки в мышцах находится определенное количество энергетических веществ (например, креатинфосфата, гликогена и др.). В результате тренировки происходит снижение уровня этих веществ в скелетных мышцах. После окончания тренировки, в фазе восстановления уровень энергетических веществ в мышечном волокне превышает исходный, то есть происходит суперкомпенсация.
Долговременная адаптация возможна только в том случае, если достигаемые срочный и отставленный тренировочные эффекты от каждой тренировки будут суммироваться (рис.1 b). Поэтому для получения определенного тренировочного эффекта и последующего повышения физической работоспособности очередную физическую нагрузку следует проводить в период преимущественно суперкомпенсации после предшествующей работы. Слишком частые (рис.1 c) тренировки прерывают стадию восстановления до достижения эффекта суперкомпенсации. Вследствие этого возможно постепенное развитие процесса недовосстановления работоспособности и преждевременное наступление утомления, что негативно сказывается на результатах. Слишком редкие тренировки (рис.1 d) в фазе сниженной суперкомпенсации не позволяют закрепить тренировочный эффект, так как каждая последующая тренировка проводится после возвращения функциональных возможностей организма к исходному уровню.
Однако еще в начале ХХ века М.Е. Маршак (1931) установил, что процессы восстановления после тяжелой мышечной работы в различных вегетативных системах протекают гетерохронно, то есть с разной скоростью. Более того, даже в пределах одной и той же системы для разных показателей ее функции возвращаются к уровню покоя не одновременно. В последующем эти данные были подтверждены для многих физиологических и биохимических показателей.
Следует отметить, что процессы восстановления энергетических веществ в мышцах также протекают с разной скоростью и завершаются в разное время. Вначале из скелетных мышц и крови удаляется молочная кислота, которая окисляется до СО2 или включается в синтез гликогена, затем происходит ресинтез креатинфосфата, гликогена и жиров.
Существует правило Энгельгардта, согласно которому интенсивность протекания восстановительных процессов и сроки восстановления энергетических запасов организма зависят от интенсивности их расходования во время выполнения упражнения. Следовательно, чем больше расход энергетических запасов мышц при работе, тем интенсивнее идет их восстановление и тем значительнее превышение исходного уровня в фазе суперкомпенсации. Однако это правило применимо лишь в ограниченных пределах. На основании эмпирических исследований установлено, что во взаимоотношениях тренировочных раздражителей и адаптационных реакций лежат следующие закономерности (Ю. Хартманн, Х. Тюнеманн, 1988):
- подпороговые раздражители (30% нагрузки от максимально возможной) не вызывают никаких адаптационных сдвигов;
- слишком высокие раздражители (большой объем и интенсивность нагрузки) вызывают спад результатов;
- оптимальные нагрузки приводят к оптимальным результатам.
В процессы адаптации, возникающие при интенсивных физических нагрузках, вовлекаются все системы, обеспечивающие функционирование мышц. Наряду с увеличением синтеза сократительных белков мышц возрастает интенсивность функционирования систем, обеспечивающих их катаболизм.
В экспериментах на животных, проведенных П.З. Гудзем (1963), в которых животные вначале подвергались различным по длительности и интенсивности физическим нагрузкам, а затем помещались в тесные клетки, было установлено следующее. Если животные получали умеренные динамические нагрузки или повышенные статические, а затем были переведены в режим гиподинамии, длительное пребывание в тесных клетках не вызвало существенных деструктивных изменений в скелетных мышцах. Однако у животных, которые до этого получали повышенные физические нагрузки, через четыре недели пребывания в тесных клетках были обнаружены дистрофические и деструктивные изменения гипертрофированных мышечных волокон. Из этого можно сделать вывод, что адаптированные к высоким физическим нагрузкам системы, отвечающие за катаболизм белка, после резкого снижения физических нагрузок продолжают функционировать некоторое время с максимальной мощностью, что приводит к резкой дистрофии и деструктивным изменениям в мышечных волокнах.
Третьим условием достижения прочных адаптационных сдвигов является постепенное увеличение физических нагрузок как по объему, так и по интенсивности. Если это условие не будет соблюдено, то по мере адаптации организма к тренировочным нагрузкам будет постепенно снижаться величина энерготрат и изменения метаболизма будут менее выраженными (М.И. Калинский, В.А. Рогозкин, 1989).
С биохимической точки зрения можно выделить несколько факторов, изменения которых существенно влияют на обмен веществ тренированного с помощью физических нагрузок организма. Во-первых, в скелетных мышцах и других органах и тканях повышаются запасы энергетических ресурсов (креатинфосфата и гликогена). Во-вторых, расширяются потенциальные возможности ферментного аппарата: повышается активность ферментов гликолиза, цикла лимонной кислоты, окисления жирных кислот, систем транспорта ионов. В-третьих, улучшаются механизмы регуляции обмена веществ с участием нервной и эндокринной систем, а также внутриклеточной системы автономного регулирования. Все эти факторы – наличие повышенного количества энергетических ресурсов и увеличенная активность ферментных комплексов, обеспечивающих основные циклы энергетического метаболизма – открывают возможности для более быстрого и более длительного пополнения запасов АТФ в организме.
В отличие от креатинфосфата и гликогена, концентрация АТФ в тканях тренированного организма не возрастает, однако, меняется скорость обмена молекул АТФ, так как повышается каталитическая активность ферментов, участвующих в гидролизе АТФ во время мышечного сокращения и в процессе ресинтеза. Под влиянием физических нагрузок в скелетных мышцах увеличивается концентрация креатинфосфата и повышается активность фермента креатинкиназы, участвующего в ресинтезе АТФ. Это приводит к расширению энергетических ресурсов в мышце и повышению скорости восстановления АТФ из креатинфосфата.
Саркоплазматическая и миофибриллярная гипертрофии скелетных мышц возможны только в том случае, если будут соблюдены описанные выше условия.
- Маршак, М.Е. О восстановительном периоде после мышечной работы / М.Е. Маршак // Физиологический журнал СССР, 1931. – Т.14. – № 2-3. – С. 204.
- Яковлев Н. Н. Очерки по биохимии спорта. М.: Физкультура и спорт, 1955. 264 с.
- Ямпольский, Л.И. Расходование и ресинтез гликогена мышц в зависимости от характера мышечной деятельности / Л.И. Ямпольский: Автореф. дис…канд. биол. наук, 1949. – 20 с.
- Хартманн, Ю. Современная силовая тренировка / Ю. Хартманн, Х. Тюнеманн. – Берлин: Шпортферлаг, 1988. – 335 с.
- Самсонова А. В. Гипертрофия скелетных мышц человека: Учеб. пособие. СПб: Кинетика, 2018. 159 с.
[2] Катаболизм – совокупность химических реакций, за счет которых крупные молекулы превращаются в молекулы меньшего размера.
[3] Анаболизм – совокупность химических реакций синтеза.
Похожие записи:
Тест времени реакции на сигнал
Представлена программа расчета времени реакции на сигнал, предназначенная для использования в учебных целях, например на занятиях по…
Саркоплазматическая гипертрофия мышц
Дано определение и описаны механизмы саркоплазматической гипертрофии скелетных мышц. Показано, что этот вид гипертрофии мышц широко…
Классификация типов конституции человека М.В. Черноруцкого
Рассмотрена классификация типов конституции человека, разработанная выдающимся терапевтом М.В. Черноруцким в 1925 году. Классификация типов конституции человека М.В.
Типы гипертрофии скелетных мышц человека
В статье дается классификация различных видов гипертрофии скелетных мышц человека на основе ряда классификационных признаков: времени проявления…
Миомейкер: Мембранный активатор слияния миобластов и образования мышц
Ученые установили, что для образования мышечных волокон необходимо слияние клеток-предшественников, которые называются миобластами. Эти клетки имеют только…
В статье изложен обзор некоторых литературных данных о механизмах работы мышц во время различных по интенсивности нагрузок. Проведён анализ адаптивности, пластичности и степени изменения мышц на разных уровнях под влиянием различных факторов. Эти изменения связаны с увеличением мускульной силы и сопровождаются трансформацией одних типов мышечных волокон в другие, изменением метаболических процессов и увеличением синтеза сократительных белков, что в основном и обуславливает процессы мышечной гипертрофии. Главным пусковым механизмом данных процессов является синтез ростовых факторов, которые выполняют, по сути, основополагающую роль. Кроме усиления синтеза белка, данные факторы способны влиять и на миосателлиты. Именно эти клетки, располагающиеся на периферии симпласта, способны делиться в постнатальном периоде. В условиях наличия нагрузки и как следствия появления развивающихся микротравм происходит их активация. Они составляют основу регенерации поврежденного мышечного участка. За счёт своего деления они формируют новые мышечные волокна. Данный процесс очень напоминает воспалительный ввиду участия провоспалительных клеток. Их цитокины также влияют на пролиферацию. Поэтому при мышечной работе происходит множество процессов, которые способствуют к непосредственному увеличению мышечного волокна путём как за счёт гипертрофии, так и гиперплазии.

3. Lin Y. et al. PGC-1? is associated with C2C12 Myoblast differentiation // Central European Journal of Biology. 2014. Т. 9. № 11. Р. 1030–1036.
4. Scharf M. et al. Mitogen-activated protein kinase-activated protein kinases 2 and 3 regulate SERCA2a expression and fiber type composition to modulate skeletal muscle and cardiomyocyte function // Molecular and cellular biology. 2013. Т. 33. № 13. Р. 2586–2602.
5. Meissner J.D. et al. The p38?/? mitogen-activated protein kinases mediate recruitment of CREB-binding protein to preserve fast myosin heavy chain IId/x gene activity in myotubes // Journal of Biological Chemistry. 2007. Т. 282. № 10. Р. 7265–7275.
6. Potthoff M.J., Olson E.N. MEF2: a central regulator of diverse developmental programs // Development. 2007. Т. 134. № 23. Р. 4131–4140.
На протяжении многих лет изучение процессов синтеза белков в скелетных мышцах при выполнении различных физических нагрузок остаётся актуальной проблемой биохимии и физиологии. Мышцы и их силовые характеристики очень важная составляющая организма каждого спортсмена, которая позволяет достигать результатов. В связи с прогрессивным развитием спорта и вовлечением большого количества людей в физическую культуру, тема здоровья спортсменов становится все более актуальной, интересной и увлекательной. Учитывая существующую сильную корреляцию между площадью поперечного сечения мышц и мышечной силой, стремление увеличить мышечную массу тела есть у каждого человека, занимающегося спортом. Кроме этого, необходимо помнить, что преобладание мышечной массы в организме благоприятно влияет на метаболические процессы.
Скелетная мышца – одна из наиболее пластичных структур в организме млекопитающих. При повышенной активности и нагрузке часто происходит увеличение её размеров, объёмов миофибриллярного аппарата, повышение сократительных возможностей (силы, мощности). Процесс прироста мышечной массы зависит от различных факторов: наследственных, конституциональных, а также пола, возраста, метаболизма, гормонального фона. Кроме того, с приобретением опыта тренировок становится все труднее увеличить мышечную массу, поэтому важно понимать и активно использовать все возможные механизмы этого процесса.
Клетки поперечно-полосатой мускулатуры отличаются от гладкомышечных миоцитов. Клетки скелетных мышц образуют многоядерный синцитий, основное вещество которого формируют миофибриллы, состоящие из толстых и тонких миофиламентов. Первый тип образуют молекулярные единицы и миозин, а второй тип содержит тропомиозин с тропонином и F-актин. Многие авторы считают скелетную мускулатуру гетерогенной системой относительно устройства и выполняемых функций, несмотря на её строгую организацию. Данное свойство помогает мышцам соответствовать возлагаемой на них функции. Так путём изменения количества саркомеров и миофибрилл обеспечивается их функциональная реорганизация [1].
Работа мышц проявляется их сокращением, которое начинается с появления очага возбуждения на нейромышечных окончаниях. Наружная мембрана деполяризуется, открываются кальциевые каналы, и концентрация кальция внутри клетки возрастает. Ионы кальция связываются с тропонином, при этом конформируется тропониновый комплекс. Участки цепей миозина связываются с актином, что сопровождается высвобождением энергии вследствие расщепления АТФ до АДФ и остатка фосфорной кислоты. Угол между лёгкой и тяжёлой цепями миозина изменяется и актиновый филамент перемещается к центру саркомера, что приводит к изменению длины мышцы, её сокращению [1, 2].
Клетки скелетных мышц подразделяются на два типа:
А) Миосателлиты – взрослые стволовые клетки мышечной ткани. Представляют собой основу для обновления мышц и прироста их массы;
Б) Миосимпласты – формируют многоядерный синцитий. Сами по себе являются мышечными тубами с миофибриллами внутри, по периферии которых располагаются ядра.
Нагрузки, оказываемые на мышцы, и само мышечное сокращение имеют некую зависимость. Предполагается, что первое будет напрямую соответствовать второму. Это достигается за счёт усиления экспрессии генов сократительных белков и энзимов обменных процессов. Мышечная активность сопровождается количественными и качественными изменениями в миоцитах того типа, которые необходимы для наиболее эффективного осуществления выполняемой работы [2].
Мышечные волокна делятся на медленные (I тип) и быстрые (II тип). Оба этих типа имеют различный состав, включающий в себя сократительные белки, ферменты энергетического обмена и внутриклеточный кальций.
Увеличение силы мышц проявляется структурными перестройками, которые затрагивают нервную и мышечные системы. Изменения в нервной системе проявляются трансформацией величины кортикальных полей, которые регулируют выполнение определённого вида движения, влиянием на синхронизацию моторных единиц и на обучение определенных мышц, отвечающих за выполнение данного вида движений. Таким образом, наибольшая активность мышц наблюдается именно тогда, когда она необходима для достижения максимального эффекта (активность мышц агонистов при одновременной пассивности антагонистов). Также наблюдается изменение частоты и устойчивости генерируемых импульсов и порога возбудимости мотонейронов. Изменения в мышечной системе могут быть связаны с гипертрофией скелетных мышц (увеличение размеров мышечного волокна) и с их гиперплазией (увеличение количества миоцитов) [3].
Но прежде чем переходить к последним двум процессам, необходимо разобраться с изменениями, происходящими в самих мышцах. В момент выполнения работы миоцит подвергается действию физических и гуморальных факторов (пассивные механические силы, гипоксемия, факторы роста, и т.д.). Они являются причиной запуска путей передачи сигнала внутри клеток, опосредуя транскрипцию и трансляцию генов, ответственных за синтез белков [2]. Изменения данных путей сопровождаются реорганизацией мышечных волокон, точнее их типов.
Одним из основных исходных сигналов является повышенная концентрация кальция внутри клетки и кальцинейрина. Кальцинейрин дефосфорилирует факторы транскрипции – NFAT (nuclear factor of activated T-cells), которые находятся в фосфорилированном состоянии [4]. Данные факторы в дефосфорилированной форме активируют гены-мишени, что способствует перестроению быстрых волокон в медленные.
По мере приспособления мышц к нагрузкам изменяются и процессы метаболизма в них. Существуют различные параметры, влияющие на формирование адаптивных механизмов в миоцитах при выполнении работы. Важнейшим является гипоксия, которая, в свою очередь активирует ферментные системы (фумараза, цитратсинтаза, ЛДГ) и запускает работу факторов транскрипции (PGC1). При недостатке кислорода происходит активация одной изоформы семейства гипоксия-индуцированных факторов (HIF; hypoxia inducible factor), которая проникает в ядро, связывается с определенным участком ДНК и активирует гены, отвечающие за гликолиз, потребление кислорода и ангиогенез, увеличивая данные процессы. Некоторые гормоны также способны влиять на экспрессию генов в мышечных клетках. Это такие гормоны, как инсулин, гормон роста, которые вместе с кортизолом запускают катаболические реакции в условиях метаболического и энергетического истощения [3].
Стоит напомнить, что мышцы не являются постоянными клетками, а заменяются в течение жизни. Пролиферация необходима для предотвращения апоптоза клеток (регулируемый процесс клеточной гибели) и поддержания массы скелетных мышц. Это осуществляется через динамический баланс между синтезом белков в мышцах и их распадом. Мышечная гипертрофия возникает тогда, когда синтез белков превышает их распад.
Что же наблюдается при гипертрофии и гиперплазии мышечного волокна? При растяжении и сокращении мышц происходит образование факторов роста IGF и MGF, которые могут действовать как паракринно, так и аутокринно. С одной стороны, их действие проявляется в увеличении синтеза сократительных белков мышечных волокон. Основным участником данного механизма является фосфорилированная PKB [5]. Её активация начинается с влияния на мышцу нагрузки, которая приводит к синтезу гена, запускающего путь IGF/PI3K. В ткани имеется несколько изоформ, некоторые из них (IGF-1 и MGF), взаимодействуя с рецепторами приводят к конформационным изменениям. Через фосфорилирование ряда рецепторов и происходит активация PKB, способствующая развитию анаболических реакций [6].
С другой же стороны, происходит усиление пролиферации миосателлитов, их митотическая активность приводит к формированию новых клеток, а также сопровождается слиянием их с имеющимися мышечными волокнами или даёт возможность формировать новые. Миосателлиты расположены между базальной мембраной и сарколеммой. Покоящиеся клетки активируются непосредственно травмированием мышцы и в ответ на это начинают активно делиться и соединяться с частями поврежденного волокна. Под влиянием тяжёлой изнурительной работы происходит также активация данных клеток из-за образования многочисленных микротравм мышечного волокна. Вследствие этого наблюдается явление подобное процессам, происходящим при воспалении. В зону повреждения активно мигрируют нейтрофилы и макрофаги, которые активируют синтез ранее упомянутых факторов роста, регулирующих пролиферацию и дифференцировку миосателлитов. Мышечная гипертрофия отличается от мышечной гиперплазии. При гипертрофии мышц, увеличиваются сократительные элементы, и межклеточный матрикс расширяется для поддержки роста. Гиперплазия приводит к увеличению количества мышечных волокон. Гипертрофия сократительных элементов может происходить путем добавления саркомеров либо последовательно или параллельно.
В отечественной литературе не утихают споры о патогенетических аспектах мышечного роста. Чаще всего гипертрофию скелетных мышц человека рассматривают как их долговременную адаптацию к физическим нагрузкам различной направленности. Но существует понятие о кратковременной гипертрофии скелетных мышц – то есть изменение объема мышцы в результате одной силовой тренировки. Спортсмены, выступающие в соревнованиях по бодибилдингу или бодифитнесу хорошо знают, что объем мышц можно немного увеличить за счет собственной крови и осмотического давления, если использовать специальный метод тренировки – пампинг.
Неоспоримым является факт увеличения объёма мышечных волокон. Это так называемая миофибриллярная гипертрофия, при которой происходит изменение объёма миофибрилл и плотность их укладки. Механизм связан с увеличением количества саркомеров в миофибриллах. Значительная роль при этом отводится активированным клеткам-сателлитам. Миогенные стволовые клетки начинают пролифелировать, а затем сливаются с существующими клетками или взаимодействуют между собой для формирования новых мышечных волокон. Этот механизм актуален при восстановлении травмированных клеток и при спортивной гипертрофии.
Существует множество данных, доказывающих идущий параллельно с этим процесс увеличения объёма несократительной части мышцы – саркоплазматическая гипертрофия. Это тонкие перестройки на биохимическом уровне клетки, а так же увеличение количества митохондрий. Многие авторы считают, что трансформации в саркоплазме повышают выносливость мышц. Ряд исследователей утверждает, что увеличение различных неконтрактильных элементов и жидкости действительно может привести к приросту мышечной массы, но без сопутствующего увеличения силы. Саркоплазматическая гипертрофия достигается специальными тренировками и часто описывается как нефункциональная. Однако ряд специалистов предполагают, что отек мышечных волокон вызывает увеличение синтеза белка и таким образом способствует росту сократительной ткани.
Эти процессы редко бывают сбалансированными и зависят от характера и интенсивности нагрузки. В скелетных мышцах при этом синтез мышечных белков преобладает над их распадом. Причиной такого метаболизма сторонники гипотезы ацидоза считают накопление молочной кислоты. С точки зрения другой теории – временная гипоксия запускает реперфузию мышц и активирует деление клеток-сателлитов. Последнее время широкое распространение получила гипотеза механического повреждения мышечных волокон. Микроразрывы сократительных белков и повреждения саркоплазмы сопровождается увеличением концентрации ионов кальция, что и стимулирует пролиферацию сателлитов.
Из этого следует, что механизмы мышечной гипертрофии известны и неоспоримы. Очень дискутабельным остается вопрос о наличии процесса гиперплазии мышц. Большинство авторов сходится во мнении, что увеличение количества мышечных волокон у человека не доказано, но при этом описывается возможность получения гиперплазии мышц в экспериментальных условиях у животных (млекопитающих и птиц). Некоторые исследователи допускают частичное увеличения числа волокон. На основании проведенного мета-анализа экспериментальных работ отмечено, что количество мышечных элементов увеличилось в экспериментах на птицах значительнее, чем при использовании в качестве подопытных млекопитающих. Примечательно также, что эффект гиперплазии наблюдался там, где использовались постоянные растяжения, а не упражнения, сочетающие его с расслаблением. Ряд исследователей (Kraemer, William J. и MacDougall J.) утверждают, что этот механизм может осуществляться под влиянием силовых тренировок. Однако доказательств увеличения мышечных волокон у людей недостаточно. Длительных исследований (более года) добровольцев и спортсменов не проводилось. Высказывается мнение, что это слишком короткий период для этого процесса. Гиперплазия подтверждается в биопсийном материале, а погрешность этого метода составляет около 10 %, что делает результат очень сомнительным.
Общее число волокон предопределяется генетически и практически не меняется в течение жизни без применения специальных стимуляторов. Российские ученые подтверждают, что вклад гиперплазии в процесс увеличения объема мышц составляет не более 5 % и, как правило, потенцирован использованием анаболических стероидов. Также гиперплазию могут вызывать блокаторы миостатина. Гормон роста при этом не вызывает гиперплазии.
Таким образом, при мышечной работе происходит множество процессов на разных уровнях. Начиная с изменений интенсивности обменных процессов и заканчивая изменениями механизмов нервной и гуморальной регуляции. Реорганизация мышц, лежащая в основе этих процессов, приводит к изменению многочисленных характеристик деятельности спортсменов.
Проанализировав все данные и изучив все возможные гипотезы, становится очевидным, что в увеличении мышечных волокон играют некую роль всё-таки два процесса. Первый – гипертрофия с ёе подвидами для сократительной и несократительной части мышцы (миофибриллярная и саркоплазматическая), которая, по мнению многих исследователей, занимает основополагающую роль. И второй это гиперплазия с её минимальным, но существенным вкладом.
Формы участия мышц в осуществлении двигательных актов весьма многообразны. Анатомическая классификация мышц (например, сгибатели и разгибатели, синергисты и антагонисты) не всегда соответствуют их функциональной роли в движениях. Так, некоторые двухсуставные мышцы в одном суставе осуществляют сгибание, а в другом — разгибание. Антагонист может возбуждаться одновременно с агонистом для обеспечения точности движения, и его участие помогает выполнять двигательную задачу. В связи с этим, учитывая функциональный аспект координации, в каждом конкретном двигательном акте целесообразно выделить основную мышцу (основной двигатель), вспомогательные мышцы (синергисты), антагонисты и стабилизаторы (мышцы, фиксирующие, не участвующие в движении суставы). Роль мышц не ограничивается генерацией силы, антагонисты и стабилизаторы часто функционируют в режиме растяжения под нагрузкой. Этот режим используется для плавного торможения движений, амортизации толчков.
На конечный результат движения влияют не только силы, развиваемые мышцами, но и силы немышечного происхождения. К ним относятся силы инерции, создаваемые массами звеньев тела, вовлекаемых в движение, а также силы реакции, возникающие в кинематических цепях при смещении любого из звеньев. Движение смещает различные звенья тела друг относительно друга и меняет конфигурацию тела, а следовательно, по ходу движения изменяются моменты упомянутых сил. Вследствие изменения суставных углов меняются и моменты мышечных сил. На ход движения влияет и масса звеньев тела; моменты сил тоже изменяются в процессе движения из-за изменения ориентации звеньев относительно вектора силы тяжести. В практической деятельности человек вступает во взаимодействие с предметами внешнего мира — различными инструментами, перемещаемыми грузами и т. д., и ему приходится преодолевать силы тяжести, упругости, трения, инерции, возникающие в процессе этого взаимодействия. Немышечные силы вмешиваются в процесс движения и делают необходимым непрерывное согласование с ними деятельности мышечного аппарата. Необходимо также нейтрализовывать действие непредвиденных помех движению, которые могут возникать во внешней среде, и оперативно исправлять допущенные в ходе реализации движения ошибки.
В связи с перечисленными особенностями скелетно-мышечной системы и условиями ее взаимодействия с внешним миром, управление движениями оказывается немыслимым без решения задачи согласования активности большого числа мышц.
В планировании, преобразовании и исполнении двигательной программы участвуют различные структуры нервной системы, организованные по иерархическому принципу. Двигательная программа может быть реализована различными способами. В простейшем случае центральная нервная система посылает заранее сформированную последовательность команд к мышцам, не подвергающуюся во время реализации никакой коррекции. В этом случае говорят о разомкнутой системе управления. Такой способ управления используется при осуществлении быстрых, так называемых баллистических движений. Чаще всего ход осуществления движения сравнивается с его планом на основе сигналов, поступающих от многочисленных рецепторов, и в реализуемую программу вносятся необходимые коррекции — это замкнутая система управления с обратными связями. Однако и такое управление имеет свои недостатки. Вследствие относительно малой скорости проведения сигналов, значительных задержек в центральном звене обратной связи и времени, необходимых для развития усилия мышцей после прихода активирующей посылки, коррекция движения по сигналу обратной связи может запаздывать. Поэтому во многих случаях целесообразно реагировать не на отклонение от плана движения, а на само внешнее возмущение еще до того, как оно успело вызвать это отклонение. Такое управление называют управлением по возмущению.
В осуществлении координации движений участвуют все отделы центральной нервной системы — от спинного мозга до коры большого мозга. У человека двигательные функции достигли наивысшей сложности в результате перехода к прямостоянию и прямохождению (что осложнило задачу поддержания равновесия), специализации передних конечностей для совершения тонких движений, использования двигательного аппарата для коммуникации (речь, письмо). В управление движениями человека включены высшие формы деятельности мозга, связанные с сознанием, что дало основание называть соответствующие движения произвольными.
Механизмы координации движений, роль тех или иных отделов ЦНС в управлении движениями изучаются чаще в опытах на животных, однако объектом исследования естественных движений является преимущественно человек, что обусловлено двумя обстоятельствами. Во-первых, человек в зависимости от задачи исследования может воспроизводить любую требуемую форму двигательной деятельности. Во-вторых, движения человека являются проявлением его поведения и трудовой деятельности и поэтому представляют особый интерес как с теоретической точки зрения вследствие их сложности и дифференцированности, так и с практической — в связи с их значением для медицины, физиологии труда, космонавтики, эргономики, физиологии спорта.
Методы исследования движений человека. Многообразие методических приемов, используемых при изучении координации движений человека, можно разделить на две группы. Одна группа методов ориентирована на получение сведений о процессах, лежащих в основе координации движений, путем регистрации внешних двигательных проявлений. Другие методы связаны с непосредственной регистрацией управляющих сигналов, поступающих к мышцам в процессе двигательной активности (электромиография), с регистрацией афферентных сигналов (микронейронография), изменений ЭЭГ, предшествующих началу движения.
Для регистрации механических параметров движения — траекторий, скорости, ускорения, развиваемой силы (механограмм) используют технику превращения неэлектрических величин в электрические с помощью различных датчиков. Так, с помощью тензо-датчиков можно непосредственно измерять и регистрировать силу, прилагаемую к тому или иному инструменту, или реакции опоры при ходьбе, с помощью резистивных датчиков на основе потенциометров — регистрировать изменения суставных углов при движении. Дифференцирование сигналов с помощью электронных дифференциаторов или ЭВМ позволяет одновременно с записью перемещения получать запись скорости и ускорения.
При анализе механических параметров движения может быть использован принцип циклографии — регистрации последовательных моментов движения путем фотосъемки движущегося человека через равные промежутки времени на неподвижную пластинку. Если на суставах или точках, соответствующих положению центров тяжести звеньев тела, укреплены светящиеся лампочки, то по такой записи — циклограмме, можно восстановить траекторию движения. В настоящее время чаще используются специализированные компьютерные системы регистрации движения с непосредственным вводом в ЭВМ изображений с двух телекамер, образующих стереопару. Решая обратную задачу механики по записям кинематических параметров (если известно распределение масс), можно рассчитать возникающие при движении силы, моменты в суставах, работу и мощность.
Движения глаз можно регистрировать электрически (электроокулография), с помощью контактных линз (внутри них располагается виток проволоки, в котором наводится напряжение при повороте витка в магнитном поле) и другими методами.
Анализ работы мышц при совершении двигательного акта осуществляется с помощью электромиографии. При электромиографическом исследовании движений обычно используют накожные электроды, укрепляемые над исследуемой мышцей. Многоканальный электромиограф дает возможность одновременно записывать электромиограммы нескольких мышц. Амплитуда электромиографического сигнала возрастает с увеличением развиваемой мышцей силы, поэтому электромиограммы в сопоставлении с регистрируемыми синхронно механограммами позволяют судить о силе сокращения мышц и о распределении их активности в последовательных фазах двигательного акта.
Набор средств изучения нейронных механизмов управления движениями на человеке включает регистрацию сухожильных рефлексов и рефлекса Гофмана (Н-рефлекс), дающих оценку уровня возбудимости двигательных нейронов спинного мозга в разные фазы движения. Позже стала возможной регистрация через кожу головы электрических потенциалов различных областей головного мозга, сопутствующих или предшествующих движению (потенциалы готовности). В последние годы разработаны методы неповреждающей стимуляции отдельных областей коры мозга человека с помощью чрезвычайно кратковременных импульсов высокого напряжения (электрическая стимуляция) или кратковременных и очень сильных магнитных полей, создаваемых индуктивными катушками, укрепленными над разными областями головы и индуцирующими электрические поля, достаточные для стимуляции мозговых структур (магнитная стимуляция).
Физиология движений изучает роль сигналов от различных рецепторов в планировании и осуществлении движений и поддержании позы. Среди них есть как простейшие — закрывание глаз, так и более сложные — использование призматических очков, смещающих или переворачивающих изображения окружающего мира, специальных систем, позволяющих вызвать у человека иллюзию движения зрительного окружения. Значение вестибулярного аппарата можно изучать в условиях его гальванической или калорической стимуляции, а также при искусственном изменении величины и направления вектора силы тяжести — на центрифуге, в условиях кратковременной или длительной невесомости. Поступающие от мышцы проприоцептивные сигналы можно менять, прикладывая к ее сухожилию вибрацию, вызывающую активацию рецепторов мышечных веретен.
С помощью описанных методов изучены такие сложные естественные двигательные акты, как ходьба, бег, рабочие и спортивные движения. Многие из этих методов используются в клинике при исследованиях нарушений двигательной функции вследствие поражения нервной системы или опорно-двигательного аппарата.
Ходьба и бег. Ходьба является наиболее распространенной формой локомоции человека (локомоция — активное перемещение в пространстве на расстояния, значительно превышающие размеры тела). Она относится к циклическим двигательным актам, при которых последовательные фазы движения периодически повторяются.
Сопоставление изменений межзвенных узлов в тазобедренном, коленном и голеностопном суставах с распределением по времени активности мышц приводит к выводу, что движение ноги в фазу переноса в значительной степени осуществляется за счет сил инерции подобно двухзвенному маятнику.
Повторяемость параметров движений в последовательных циклах при ходьбе не абсолютная: движения обладают некоторой вариабельностью. Наименьшая вариабельность у кинематической картины ходьбы, наибольшая — в работе мышц, проявляющаяся в изменениях электромиограмм от цикла к циклу. Это отражает корригирующую деятельность ЦНС, которая при каждом шаге вносит в стандартную структуру ходьбы поправки, необходимые для обеспечения относительного постоянства ее кинематики.
Бег отличается от ходьбы тем, что нога, которая находится позади, отталкивается от опоры раньше, чем другая опускается на эту опору. В результате в беге имеется безопорный период — период полета. В беге благодаря большим, чем при ходьбе, скоростям перемещения более значительную роль играют баллистические компоненты движения — перемещение звеньев ноги по инерции.
Рабочие движения. Рабочими движениями в широком смысле слова могут быть названы самые разные целенаправленные движения, совершаемые как в процессе труда, так и в повседневной жизни.
У человека основным рабочим органом является рука, причем для выполнения двигательной задачи обычно наиболее важно положение кисти, которая в результате движения должна в определенный момент оказаться в определенном месте пространства. Благодаря большому числу степеней свободы верхней конечности кисть может попасть в нужную точку по разным траекториям и при различных соотношениях углов в плечевом, локтевом и лучезапястном суставах. Это многообразие возможностей позволяет выполнять двигательную задачу, начиная движение из различных исходных поз, и в то же время ставит ЦНС перед сложной задачей выбора одного варианта из многих.
Электромиографические исследования ряда рабочих движений показали сложную картину работы мышц, однако в этой картине часто можно выделить устойчивые сочетания активности некоторых мышц, используемые в различных движениях. Это синергии, основанные на врожденных или выработанных в процессе опыта связях, которые, являясь устойчивыми компонентами движений, упрощают управление сложными двигательными актами и помогают преодолеть избыточность числа мышц и количества степеней свободы.
При совершении одного и того же, даже простого, движения организация мышечной деятельности в сильной степени зависит от вмешательства немышечных сил, в частности, внешних по отношению к человеку. Так, при ударе молотком, когда к массе предплечья добавляется масса молотка, и, следовательно, возрастает роль инерции, разгибание предплечья совершается по типу баллистического движения — мышцы-разгибатели активны только в начале разгибания, которое дальше совершается по инерции, а в конце притормаживается мышцами-антагонистами. Аналогичное по кинематике движение при работе напильником (рис. 4.19), когда основная внешняя сила — трение, совершается путем непрерывной активности мышцы на протяжении всего разгибания. Если первое из этих двух движений является в основном предпрограммированным, то во втором велика роль обратных связей.
Поза стояния у человека энергетически относительно экономна, так как моменты силы тяжести невелики вследствие близости проекции тяжести тела к осям основных суставов ног. Менее экономно стояние у многих животных, например у кошки и собаки, которые стоят на полусогнутых конечностях.
Другой важный компонент тонуса — рефлекторный, определяется рефлексом на растяжение. При исследовании на человеке он выявляется по сопротивлению растяжения мышцы в случае пассивного поворота звена конечности в суставе. Если в процессе такого исследования записать электромиограмму, то в растягиваемой мышце регистрируется электрическая активность, свидетельствующая об активации двигательных единиц. У здорового человека рефлекс на растяжение при пассивном движении наблюдается только в процессе самого растяжения и притом в случае достаточно большой скорости растяжения. Тонический компонент рефлекса на растяжение, т. е. активность в растянутой мышце, обычно отсутствует, о чем можно судить по отсутствию электромиографического сигнала.
Выработка двигательных навыков. Совершенствование двигательной функции человека в процессе онтогенеза происходит как вследствие продолжающегося в первые годы после рождения созревания отделов нервной системы и врожденных механизмов, участвующих в координации движений, так и в результате обучения, т. е. формирования новых связей, ложащихся в основу программ тех или иных конкретных двигательных актов. Координация новых, непривычных движений имеет характерные черты, отличающие ее от координации тех же движений после обучения.
Обилие степеней свободы в опорно-двигательном аппарате, влияние на результат движения сил тяжести и инерции осложняют выполнение любой двигательной задачи. На первых порах обучения нервная система справляется с этими трудностями, нейтрализуя помехи путем развития дополнительных мышечных напряжений. Мышечный аппарат жестко фиксирует суставы, не участвующие в движении, активно тормозит инерцию быстрых движений. Такой путь преодоления помех, возникающих в ходе движения, энергетически невыгоден и утомителен. Использование обратных связей еще несовершенно — коррекционные посылки, возникающие на их основе, несоразмерны и вызывают необходимость повторных дополнительных коррекций.
На электромиограммах видно, что мышцы-антагонисты даже тех суставов, в которых совершаются движения, активируются одновременно, при этом в циклических движениях мышцы почти не расслабляются. Возбуждены также многие мышцы, не имеющие прямого отношения к данному двигательному акту. Движения, совершаемые в таких условиях, напряжены и неэстетичны (например, движения человека, впервые вышедшего на коньках на лед).
Как показал в своих исследованиях Н. А. Бернштейн, по мере обучения вырабатывается такая структура двигательного акта, при которой немышечные силы включаются в его динамику, становятся составной частью двигательной программы. Излишние мышечные напряжения при этом устраняются, движение становится более устойчивым к внешним возмущениям. На электромиограммах видна концентрация возбуждения мышц во времени и пространстве, периоды активности работающих мышц укорачиваются, а количество мышц, вовлеченных в возбуждение, уменьшается. Это приводит к повышению экономичности мышечной деятельности, а движения делаются более плавными, точными и непринужденными (см. рис. 4.19).
Важную роль в обучении движениям играет рецепция, особенно проприорецепция. В процессе двигательного обучения обратные связи используются не только для коррекции движения по его ходу, но и для коррекции программы следующего движения на основе ошибок предыдущего.
Утомление. При длительной физической работе наступает утомление, которое, в частности, проявляется в изменении координации мышечной деятельности. Возбуждение каждой работающей мышцы становится менее локализованным во времени. В работу вовлекаются другие мышцы, сначала синергисты, компенсирующие снижение силы основных мышц, а затем, по мере нарастания дискоординации — и другие мышцы, в частности антагонисты. Движения становятся менее точными, темп их замедляется.
Картина мышечной активности во время движений, совершаемых на фоне утомления, во многом напоминает картину, наблюдаемую при выполнении новых, непривычных движений.
Читайте также: