
Транспортная система клетки осуществляют транспорт белков что это
Обновлено: 19.05.2024
Мембранный транспортный белок (или просто переносчик ) представляет собой мембранный белок участвует в движении ионов , малых молекул и макромолекул , таких как другой белок , через биологическую мембрану . Транспортные белки - это интегральные трансмембранные белки ; то есть они постоянно существуют внутри и охватывают мембрану, через которую они переносят вещества. Белки могут способствовать перемещению веществ за счет облегченной диффузии или активного транспорта . Два основных типа белков, участвующих в таком транспорте, в широком смысле классифицируются как каналы или носители . В растворенных носителях и атипичный ТСП являются вторичными активными или стимулированием транспортеров в организме человека. В совокупности мембранные транспортеры и каналы являются транспортомами. Транспортомы регулируют клеточный приток и отток не только ионов и питательных веществ, но и лекарств.
СОДЕРЖАНИЕ
Разница между каналами и операторами связи
Носитель не открыт одновременно как внеклеточные и внутриклеточным среды. Либо его внутренние ворота открыты, либо открыты внешние ворота. Напротив, канал может быть открыт для обеих сред одновременно, позволяя молекулам диффундировать без перерыва. Носители имеют сайты связывания, а поры и каналы - нет. Когда канал открыт, миллионы ионов могут проходить через мембрану в секунду, но обычно только от 100 до 1000 молекул проходят через молекулу-носитель за одно и то же время. Каждый белок-носитель предназначен для распознавания только одного вещества или одной группы очень похожих веществ. Исследования коррелировали дефекты в определенных белках-носителях с конкретными заболеваниями.
Активный транспорт
Действие натрий-калиевого насоса является примером первичного активного транспорта. Два белка-носителя слева используют АТФ для вывода натрия из клетки против градиента концентрации. Белки справа используют вторичный активный транспорт для перемещения калия в клетку.
Активный транспорт - это движение вещества через мембрану против градиента его концентрации. Обычно это делается для накопления высоких концентраций молекул, в которых нуждается клетка, таких как глюкоза или аминокислоты. Если в процессе используется химическая энергия, такая как аденозинтрифосфат (АТФ), это называется первичным активным транспортом . Вторичный активный транспорт включает использование электрохимического градиента и не использует энергию, производимую в ячейке. В отличие от канальных белков, которые только пассивно переносят вещества через мембраны, белки-носители могут переносить ионы и молекулы либо пассивно, за счет облегченной диффузии, либо посредством вторичного активного транспорта. Белок-носитель необходим для перемещения частиц из областей с низкой концентрацией в области с высокой концентрацией. Эти белки-носители имеют рецепторы, которые связываются со специфической молекулой (субстратом), нуждающейся в транспорте. Транспортируемая молекула или ион (субстрат) должны сначала связываться в сайте связывания в молекуле-носителе с определенным сродством связывания. После связывания и пока сайт связывания обращен в одну сторону, носитель захватит или закроет (захватит и удержит) субстрат в своей молекулярной структуре и вызовет внутреннюю транслокацию, так что отверстие в белке теперь обращено к другой стороне плазматическая мембрана. Субстрат белка-носителя высвобождается в этом месте в соответствии с его аффинностью связывания там.
Облегченная диффузия
Облегченная диффузия в клеточной мембране, показаны ионные каналы (слева) и белки-носители (три справа).
Облегченная диффузия - это прохождение молекул или ионов через биологическую мембрану через определенные транспортные белки и не требует затрат энергии. Облегченная диффузия используется особенно в случае больших полярных молекул и заряженных ионов; как только такие ионы растворяются в воде, они не могут свободно диффундировать через клеточные мембраны из-за гидрофобной природы жирнокислотных хвостов фосфолипидов, составляющих бислои. Тип белков-носителей, используемых для облегченной диффузии, немного отличается от тех, которые используются для активного транспорта. Они по-прежнему являются трансмембранными белками-переносчиками, но это закрытые трансмембранные каналы, что означает, что они не перемещаются внутри и не требуют АТФ для функционирования. Субстрат берется с одной стороны закрытого носителя, и без использования АТФ субстрат высвобождается в клетку. Их можно использовать как потенциальные биомаркеры.
Обратная диффузия
Обратный транспорт или реверсирование переносчика - это явление, при котором субстраты мембранного транспортного белка перемещаются в направлении, противоположном их типичному перемещению переносчиком. Обращение переносчика обычно происходит, когда белок мембранного транспорта фосфорилируется определенной протеинкиназой , которая представляет собой фермент, который добавляет фосфатную группу к белкам.
(Сгруппированы по категориям базы данных классификации транспортеров)
1: каналы / поры
- α-спиральные белковые каналы, такие как потенциал-управляемые ионные каналы (VIC), лиганд-управляемые ионные каналы (LGIC)
- β-бочкообразные порины, такие как аквапорин
- каналообразующие токсины, в том числе колицины , дифтерийный токсин и другие
- Каналы, не синтезируемые рибосомами, такие как грамицидин
- Холинс ; которые участвуют в экспорте ферментов, которые переваривают стенки бактериальных клеток на ранней стадии лизиса клеток.
Облегченная диффузия происходит в клеточную мембрану и из нее через каналы / поры и носители / переносчики.
Каналы либо в открытом, либо в закрытом состоянии. Когда канал открывается с небольшим конформационным переключением, он открыт для обеих сред одновременно (внеклеточной и внутриклеточной).

На этой картинке изображен симпорт. Желтый треугольник показывает градиент концентрации для желтых кругов, в то время как зеленый треугольник показывает градиент концентрации для зеленых кругов, а пурпурные стержни представляют собой связку транспортного белка. Зеленые круги движутся против своего градиента концентрации через транспортный белок, который требует энергии, в то время как желтые круги движутся вниз по градиенту концентрации, который высвобождает энергию. Желтые круги производят больше энергии за счет хемиосмоса, чем требуется для перемещения зеленых кругов, поэтому движение связано, а некоторая энергия нейтрализуется. Одним из примеров является пермеаза лактозы, которая позволяет протонам снижать градиент концентрации в клетке, а также закачивать в клетку лактозу.
Поры постоянно открыты для этих обеих сред, потому что они не претерпевают конформационных изменений. Они всегда открыты и активны.
2: Электрохимические транспортеры, управляемые потенциалом
Также называются белками-носителями или вторичными носителями.

На картинке изображен унипорт. Желтый треугольник показывает градиент концентрации для желтых кружков, а пурпурные стержни - пучок транспортных белков. Поскольку они движутся вниз по градиенту концентрации через транспортный белок, они могут выделять энергию в результате хемиосмоса . Одним из примеров является GLUT1, который перемещает глюкозу вниз по градиенту ее концентрации в клетку.
- Переносчик дофамина (DAT)
- Транспортер норэпинефрина (NET)
- Транспортер серотонина (SERT)
- Транспортеры везикулярных моноаминов (VMAT)
- Нигерицина семьи
- Ionomycin семьи
3: Основные активные транспортеры

На этой картинке изображен антипорт. Желтый треугольник показывает градиент концентрации для желтых кружков, в то время как синий треугольник показывает градиент концентрации для синих кружков, а пурпурные стержни представляют собой связку транспортного белка. Синие кружки движутся против своего градиента концентрации через транспортный белок, который требует энергии, в то время как желтые кружки движутся вниз по градиенту концентрации, который высвобождает энергию. Желтые круги производят больше энергии за счет хемиосмоса, чем требуется для перемещения синих кругов, поэтому движение связано, а некоторая энергия нейтрализуется. Одним из примеров является натрий-протонный обменник, который позволяет протонам снижаться по градиенту их концентрации в ячейке, одновременно откачивая натрий из ячейки.
4: Групповые транслокаторы
Групповые транслокаторы обеспечивают особый механизм фосфорилирования сахаров, когда они транспортируются в бактерии (групповая транслокация PEP).
5: Электронные носители
Трансмембранные переносчики электронов в мембране включают двухэлектронные переносчики, такие как оксидоредуктазы с дисульфидной связью (DsbB и DsbD в E. coli), а также одноэлектронные переносчики, такие как НАДФН-оксидаза. Часто эти окислительно-восстановительные белки не считаются транспортными белками.
Примеры
Каждый белок-носитель, особенно внутри одной и той же клеточной мембраны, специфичен для одного типа или семейства молекул. Например, GLUT1 - это названный белок-носитель, обнаруженный почти во всех мембранах клеток животных, который переносит глюкозу через бислой. Другие специфические белки-носители также важными способами помогают организму функционировать. Цитохромы действуют в цепи переноса электронов как белки-переносчики электронов.
Патология
Ряд наследственных заболеваний включает дефекты белков-носителей в определенном веществе или группе клеток. Цистеинурия (цистеин в моче и мочевом пузыре) - это заболевание, связанное с дефектными белками-переносчиками цистеина в мембранах клеток почек. Эта транспортная система обычно удаляет цистеин из жидкости, которая должна стать мочой, и возвращает эту незаменимую аминокислоту в кровь. Когда этот носитель не работает, большие количества цистеина остаются в моче, где он относительно нерастворим и имеет тенденцию выпадать в осадок. Это одна из причин мочевых камней. Было показано, что некоторые белки-переносчики витаминов сверхэкспрессируются у пациентов со злокачественными заболеваниями. Например, было показано , что уровни белка-носителя рибофлавина (RCP) значительно повышены у людей с раком груди .
Смотрите также
использованная литература
Андерле, П., Барбачору, Ч., Бусси, К., Дай, З., Хуанг, Ю., Папп, А., Рейнхольд, В., Сади, В., Шанкаварам, У., и Вайнштейн, Дж. (2004). Мембранные транспортеры и каналы: роль транспортома в химиочувствительности рака и химиорезистентности. Исследования рака, 54, 4294-4301.
Переносчики и каналы — устройства, обеспечивающие специфические транспортные процессы. В клетках высших организмов, и в том числе в кишечных, существуют транспортеры многих типов. К ним относятся переносчики глюкозы, аминокислот и др. Каждое такое устройство переносит один или ограниченное число типов органических молекул через мембрану по электрохимическому градиенту или благодаря сопряжению с механизмом транспорта другого вещества, движение которого до градиенту концентраций служит источником энергии для сопряженного с ним процесса.
Для вторичной энергизации используются многие ионные градиенты, но преимущественно градиент Na + без участия АТФ (табл. 8). Na + -зависимые переносчики используются в различных системах, включающих в себя систему всасывания (например, тонкая кишка), систему обеспечения собственных энергетических и пластических потребностей, транспорт ряда веществ (например, тех же глюкозы и аминокислот) для собственных синтезов и т.д. В печени механизмы активного транспорта глюкозы используются для депондрования веществ и их иммобилизации из депо. Мобильный переносчик, представляющий собой, как правило, белковую молекулу, движется от одной поверхности мембраны к другой, совершая вертикальные или вращательные движения, с тем чтобы связывать транспортируемые субстраты на одной поверхности мембраны и освобождать с другой. Типичным примером такого переносчика служат ионофоры. Канал как устройство характеризуется наличием постоянной или индуцированной поры, через которую проходит транспортируемое вещество.
Nа + -зависимый транспорт органических веществ в клетках животных
Ткани и клетки, для которых характерен Na + -завксимын транспорт аминокислот
Эпителиальные ткани Тонкая кишка (животные различных видов) Почки Желчный пузырь (кролик) Мочевой пузырь земноводных Стенка тела (морские беспозвоночные) Неэпителиальные ткани Жировые ткани Хрусталик глаза Кости и хрящи Печень Мозг Поперечнополосатые мышцы Одиночные клетки Асцитная карцинома Эрлиха Фибробласты мыши Морские бактерии Ретикулоциты (кролик) Эритроциты голубя Лейкоциты (человек)
Вместе с тем, как упомянуто выше, наряду с Na + -зависимым транспортом существует Ма + -пезависимый транспорт. Этот тип транспорта обнаружен при изучении переноса через мембрану кишечных клеток моносахаридов, аминокислот и других мономеров, образующихся в результате мембранного гидролиза соответствующих олигомеров.
Таблица 10
Некоторые примеры генетических нарушений транспортных систем у человека
Благодаря распространению Na + -зависимых транспортеров на апикальной мембране, а натриевых насосов — на базолатеральной создается транзитный перенос, необходимый для обеспечения за счет работы каждой кишечной клетки потребностей многих тысяч других соматических клеток. Транспортные функции клеток других типов реализуются практически тем же самым набором функциональных блоков. Действительно, транспортные характеристики клеток тонкой кишки и плаценты во многом совпадают.
Ряд клеток характеризуется тем, что универсальные транспортные блоки расположены на клеточной поверхности иначе. В частности, в ацинарных клетках поджелудочной железы, осуществляющих внешнюю секрецию ферментов, насосы и Na + -зависимые транспортеры расположены в базолатеральной мембране. Этого достаточно для обеспечения градиента Na + между вне- и внутриклеточной жидкостями и поступлением нутриентов из крови в цитоплазму, а также для энергизации многих других процессов. Кроме того, локализация насосов и каналов в базолатеральной мембране исключает интенсивное поступление потоков воды и солей в апикально-базальном направлении.
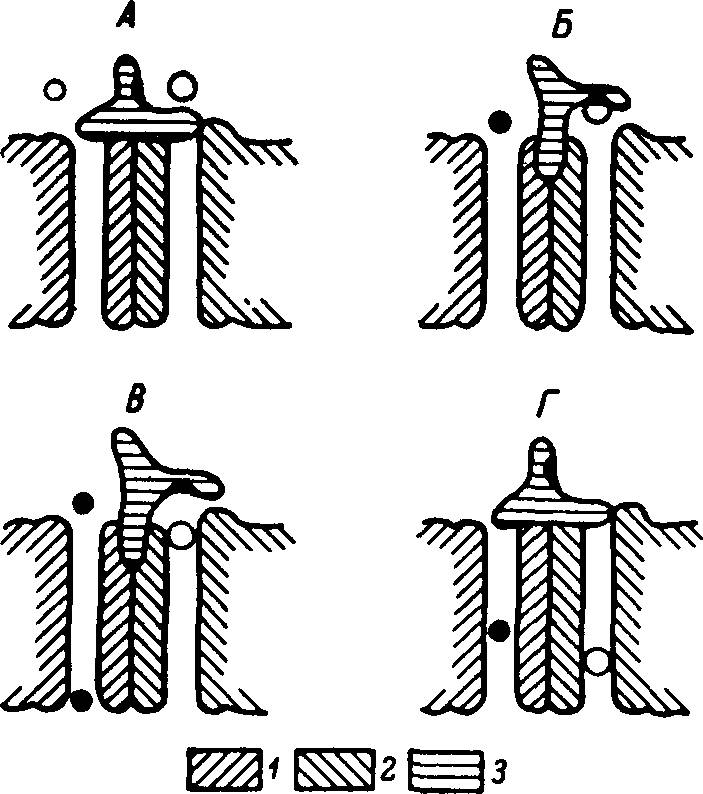
Рис. 28. Схема действия транспортера глюкозы и натрия в плазматической мембране.
А — исходное состояние транспортера: оба канала закрыты; Б — после связывания глюкозы с аллостерическим центром на воротном устройстве: натриевый канал открылся; В — при прохождении натрия по натриевому каналу произошла аллостерическая активация глюкозного канала и молекула глюкозы перешла с воротного устройства на вход своего канала; Г — освобождение центра на воротном устройстве влечет за собой закрытие натриевого и глюкозного каналов. 1 — натриевый канал; 2 — глюкозный канал; 3 — воротное устройство. Светлые пружки — молекулы глюкозы, черные — ионы натрия.
Согласно современным представлениям, при Na + -зависимом транспорте глюкозы через плазматическую мембрану клеток эукариотов происходит образование тройного комплекса: Na + —глюкоза—транспортер. Однако более адекватной представляется наша модель транспортера (рис. 28). Она характеризуется наличием двух параллельно взаимодействующих каналов (для Na + и для глюкозы) и поверхностного (воротного) белка, связывающего глюкозу на входе в транспортную систему. Транспортный цикл такой модели состоит в следующем: 1) исходно глюкозный и натриевый каналы неактивны; 2) при связывании глюкозы с аллостерическим центром на воротном белке натриевый канал активируется (эта стадия напоминает контроль натриевой проницаемости ацетилхолином) и Na + движется из экстра- в интрацеллюлярную жидкость; 3) на определенном этапе движения Na + по каналу происходит аллостерическая активация глюкозного канала; 4) через активированный канал транспортируется молекула глюкозы, первоначально фиксированная в воротном устройстве; 5) освобождение воротного устройства сопровождается дезактивацией натриевого канала. Реактивация последнего происходит при связывании с аллостерическим центром следующей молекулы глюкозы. При температуре 16° С глюкозный канал инактивируется, а натриевый продолжает функционировать. Из нашей модели вытекает, что скорость транспорта глюкозы и Na + в определенном диапазоне зависит от концентрации Na + во внеклеточной жидкости. Важно, что Ка + -зависимые транспортеры глюкозы и аминокислот, поскольку показана их идентичность в клетках различных типов и у организмов разных групп, должны соответствовать одной из моделей молекулярных машин.
Несмотря на то что сравнительная физиология ионных каналов изучена недостаточно, существуют данные о значительном сходстве их молекулярной структуры и функциональной организации. Вероятно, у млекопитающих и насекомых имеются все основные типы ионных каналов — натриевые, калиевые, хлорные и др.
Далее, у млекопитающих, рыб и насекомых продемонстрировано наличие специализированных натриевых каналов, чувствительных к амилориду, а также каналов, реализующих котранспорт Na + и Сl - , чувствительных к фуросемиду. Интересно, что те же самые ингибиторы специфически тормозят транспорт ионов в мальпигиевых канальцах и толстой кишке насекомых. Недавно выявлен котранспортер, осуществляющий котранспорт в клетку одного иона Na + , одного иона К + и двух ионов Сl - . Этот механизм обнаружен в клетках почки и тонкой кишки млекопитающих, в клетках различных органов рыб и насекомых.
До недавнего времени считалось, что функции секреции, экскреции и всасывания имеют глубокие различия, касающиеся механизмов функционирования и эволюционного происхождения. Однако между этими процессами принципиальной разницы не существует. Все основные функциональные блоки, упомянутые выше, лежат и в основе перечисленных функций, в ряде случаев полярных по своему физиологическому значению (например, всасывание и экскреция).
Характеристики ферментных и транспортных систем клеток, выполняющих различные функции (например, всасывание в тонкой кишке, реабсорбция воды в протоках пищеварительных желез и т.д.), очень близки. В апикальной мембране клеток тонкой кишки и почечных канальцев присутствуют трансмембранные ферменты, Ка + -зависимые транспортеры глюкозы, аминокислот и других нутриентов. В базолатеральной мембране этих клеток локализованы близкие или идентичные по молекулярной структуре АТФазы. Системы с множеством транспортеров, локализованных в апикальной мембране, и натриевых насосов, расположенных в базолатеральной мембране, распространены очень широко. В некоторых структурах они выполняют не только резорбтивную, но и секреторную функцию.
Существенно, что при электронной микроскопии многие субклеточные и молекулярные структуры клеток тонкой кишки человека и высших животных неотличимы от таковых клеток кишки рыб, насекомых, моллюсков, круглых червей и т.д., а клетки тонкой кишки — от клеток плаценты, канальцев почек и других клеток.
1. Детерминанты половых клеток
1. Детерминанты половых клеток Будущие половые клетки — гоноциты иногда можно отличить уже на очень ранних стадиях развития. Классическим примером является аскарида, у которой после первого деления яйца в одном из двух бластомеров теряются концевые участки хромосом и
2. Пол половых клеток
2. Пол половых клеток После того как гоноциты попали в половые железы, их судьба зависит от того, какие половые клетки они должны образовать — мужские, т. е. сперматозоиды, или женские, т. е. яйца. В первом случае их путь развития называют сперматогенезом, а во втором —
1. Форма клеток
1. Форма клеток Форма клеток зависит от их внутренней структуры и свойств клеточной оболочки и от их окружения — соседних клеток и поверхностей контакта. Так, при культивировании отдельных клеток на поверхности стекла все клетки стремятся распластаться по субстрату.
3. Перемещения отдельных клеток
3. Перемещения отдельных клеток Многие процессы формообразования происходят не путем перемещения клеточных пластов, а путем перемещения отдельных клеток. Иногда движение клеточного пласта происходит при частичной потере его эпителиальной структуры. Так, например,
Два главных органа иммунной системы вырабатывают два сорта клеток: Т–и В–лимфоциты.
Два главных органа иммунной системы вырабатывают два сорта клеток: Т–и В–лимфоциты. — Следовательно, иммунная система каждого организма имеет два главных органа, не так ли? — Так? — Один, тимус, заведует выработкой лимфоцитов, способных превращаться в
§ 8. Заряды мембраны нервных клеток
§ 8. Заряды мембраны нервных клеток Однако основные свойства нервной системы обусловлены способностью быстро реагировать на изменение ситуации внутри или вне организма. Скоростные процессы не могут осуществляться по медленным гуморальным законам, они происходят по
7.8. Сигналы нервных клеток
7.8. Сигналы нервных клеток Нервная система возникает в ходе эволюции для анализа организмами поступающей информации. С этой целью нервные клетки используют электрические и химические сигналы. Электрические сигналы бывают двух видов.Градуальный потенциал возникает в
5.3. Транспортные системы клеток. Насосы
5.3. Транспортные системы клеток. Насосы Под насосами подразумеваются системы, в которых сочетаются механизмы энергизации и трансмембранного переноса. Принимается, что источником энергии в большинстве случаев служит энергия макроэргических связей АТФ. Деятельность
2.7. Попытки моделирования примитивных клеток
2.7. Попытки моделирования примитивных клеток В ранних работах о происхождении жизни была выдвинута идея предклеточных структур, существующих в водной среде в форме “микросфер” (Fox, 1965; 1991) и “коацерватных капель” (Опарин, 1966). Микросферы формировались из т. н.
9.3. Возникновение клеток
9.3. Возникновение клеток Предполагается, что этапом возникновения жизни на Земле следует считать период, когда сформировались простейшие клеточные системы, ставшие элементарной ячейкой живого. Сведения, касающиеся этой проблемы, освещены в обзорах, посвященных
9.3. Возникновение клеток
9.3. Возникновение клеток Предполагается, что этапом возникновения жизни на Земле следует считать период, когда сформировались простейшие клеточные системы, ставшие элементарной ячейкой живого. Сведения, касающиеся этой проблемы, освещены в обзорах, посвященных

Новость
Транспортировка молекул внутри мембранных пузырьков (везикул) — основная разновидность внутри- и внеклеточного транспорта у эукариот
Автор
Редакторы

Гены везикулярного транспорта: при чем тут дрожжи?
Рэнди Шекман (Randy W. Schekman), изучавший биохимию под началом Артура Корнберга (лауреата Нобелевской премии в 1959 году), решил заняться механизмом молекулярного транспорта с позиций генетики и выбрал в качестве модельного объекта пекарские дрожжи (Saccharomyces cerevisiae). Шекман собрался выявить гены, которые участвуют в процессе везикулярного транспорта, с помощью мутантных форм дрожжей и анализа накапливающихся внутри клетки секреторных ферментов [1–3].
Работы Шекмана заложили генетический базис и выявили ключевые регуляторные события в везикулярном транспорте.

Ключевые белки везикулярного транспорта
Следующим стало открытие фактора SNAP (soluble NSF-attachment protein), связывающегося с мембраной при участии NSF [12]. Знаковым событием, соединившим работы Шекмана и Ротмана, стало понимание того, что ген sec18 кодировал как раз NSF, а sec17 — SNAP [13–15] (потом было найдено соответствие и остальных генов прочим открытым Ротманом белкам). Это открытие показало, что система везикулярного транспорта является эволюционно древней для эукариот.
Ротман разобрался в работе везикулярного транспорта и слиянии мембран, а также в биохимических экспериментах показал, каким образом реализуется специфичность этих процессов.

Временнóй контроль слияния везикул
Томас Зюдоф (Thomas C. Südhof) исходно работал в Германии, а потом переехал в США. В организованной им научной группе решали задачу регуляции слияния мембранных везикул, поскольку было понятно, что это событие должно запускаться каким-то внешним сигналом, а вот каким — никто не знал. Эта задача была подсказана процессами выброса медиаторов в синапсах или секреции инсулина в поджелудочной железе, где требуется очень точный временнóй контроль.
Зюдоф заметил, что выброс медиаторов управляется внутриклеточной концентрацией ионов кальция (Ca 2+ ) вблизи пресинаптической мембраны. В итоге он открыл, что комплексин и синаптотагмин являются критически важными компонентами кальций-зависимого слияния мембран (рис. 4).

Зюдоф внес решающий вклад в нейробиологию и клеточную биологию, показав, каким образом осуществляется временной контроль выброса медиаторов в синапсах, а также установив роль ионов кальция в этом процессе.
Везикулярный транспорт и медицина
Работы Ротмана, Шекмана и Зюдофа приподняли завесу тайны над клеточным транспортом в клетке и показали, каким образом молекулы своевременно доставляются в нужное место. Несложно догадаться, что нарушения везикулярного транспорта, учитывая его роль в жизни клетки, приводят к серьезным болезням как нервной, так и эндокринной систем.
Например, такие метаболические расстройства как диабет второго типа характеризуются нарушениями как в секреции инсулина, так и в инсулинзависимом транспорте глюкозы в мышечной и жировой тканях. Кроме того, клетки иммунитета используют везикулярный транспорт для секреции цитокинов и прочих иммунологических молекул, управляющих врожденным и приобретенным иммунитетом.
Учитывая вышесказанное, открытия Ротмана, Шекмана и Зюдофа могут пролить свет на механизмы этих заболеваний и дать ключ к их лечению.
Транспортные белки – это белки, которые транспортируют вещества через биологические мембраны. Транспортные белки находятся внутри самой мембраны, где они образуют канал или несущий механизм, чтобы позволить их подложка переходить с одной стороны на другую.
Вещества, переносимые этими белками, могут включать ионы, такие как натрий и калий; сахара, такие как глюкоза; белки и молекулы-мессенджеры; и многое другое.
Транспортные белки обычно осуществляют два вида транспорта:облегченная диффузия ”, Где транспортный белок просто создает отверстие для вещества, чтобы распространять вниз его градиент концентрации ; а также “активный транспорт,” где клетка расходует энергию, чтобы переместить вещество против градиента его концентрации.
Функция транспортного белка
Жизнь, как мы знаем, зависит от способности клеток избирательно перемещать вещества, когда им это необходимо. Определенные важные молекулы, такие как ДНК, должны постоянно находиться внутри клетки; но другие молекулы, такие как ионы, сахара и белки, возможно, должны пройти внутрь и наружу, чтобы клетка функционировала должным образом.
Перемещая вещества через мембраны, транспортные белки делают возможным все – от нервных импульсов до клеточного метаболизма.
Без транспортных белков, например, градиент натрия-калия, который позволяет нашим нервам срабатывать, не существовал бы.
Типы транспортных белков
Каналы / Поры
Эти белки характеризуются тем, что они открыты как для внутриклеточного, так и для внеклеточного пространства одновременно. Напротив, белки-носители открыты только внутри или снаружи клетки в любой момент времени.
Каналы или поры обычно сконструированы так, что через них может проходить только одно конкретное вещество.
Например, в ионно-управляемых каналах напряжения часто используются заряженные аминокислоты, расположенные на точных расстояниях, чтобы привлечь их желаемый ион, отталкивая все остальные. Затем желаемый ион может течь через канал, тогда как другие вещества не могут.
Ионные каналы, управляемые напряжением, являются хорошими примерами транспортных белков, которые действуют по мере необходимости. Часто обнаруживаемые в нейронах, управляемые напряжением ионные каналы открываются в ответ на изменения электрохимического потенциала мембраны.
В закрытом состоянии управляемый напряжением канал не позволяет ионам проходить через клеточная мембрана, Но когда он открыт, он позволяет огромным количествам ионов проходить очень быстро, позволяя клетке быстро менять свой мембранный потенциал и запускать нервный импульс.
Белки-носители
Белки-носители представляют собой транспортные белки, которые открыты только с одной стороны мембраны одновременно.
Они часто проектируются таким образом, потому что они транспортируют вещества против градиента их концентрации. Одновременное открывание по обе стороны мембраны может позволить этим веществам просто течь обратно вдоль градиента концентрации, сводя на нет белок-носитель Работа
Для выполнения своей работы белки-носители обычно используют энергию для изменения формы.
Например, натриево-калиевая помпа использует энергию АТФ, чтобы изменить свою форму с открытой на внутриклеточную. решение, чтобы быть открытым для внеклеточного раствора. Это позволяет ему собирать ионы внутри клетки и выпускать их снаружи, а затем наоборот.
Другие белки-носители могут использовать другие источники энергии, такие как существующие градиенты концентрации, для достижения «вторичного активный транспорт «. Это означает, что их транспорт стал возможен благодаря энергии, расходуемой клеткой, но сам белок не использует АТФ напрямую.
Хорошим примером является транспортный белок натрий-глюкоза, который использует градиент концентрации натрия, первоначально созданный натриево-калиевым насосом, для перемещения глюкозы против градиента концентрации.
Мы обсуждаем натрий-калиевый насос и натрий-глюкозный транспортный белок подробно ниже.
Примеры транспортных белков
Натриево-калиевый насос
Наиболее известным примером первичного активного транспортного белка является натриево-калиевая помпа. Именно этот насос создает ионный градиент, который позволяет нейронам срабатывать.
Натриево-калиевая помпа начинается с натриевых сайтов связывания, обращенных внутрь клетки. Эти сайты привлекают ионы натрия и удерживают их.
Когда каждый из трех сайтов связывания натрия связывает ион натрия, белок связывается с молекула АТФ, и разбивает его на ADP + фосфатная группа, Белок использует энергию, выделяемую в этом процессе, чтобы изменить форму.
Теперь сайты связывания натрия сталкиваются с внеклеточным раствором. Они выделяют три иона натрия вне клетки, в то время как сайты связывания белка с калием связываются с двумя ионами калия.
Когда оба калий-связывающих сайта заполнены, белок возвращается к своей первоначальной форме. Теперь ионы калия высвобождаются внутри клетки, и пустые сайты связывания натрия могут связывать больше ионов натрия.

Когда наступает время для срабатывания нервной клетки, сильные электрические и химические градиенты позволяют клетке производить огромное мгновенное изменение, открывая свои управляемые напряжением ионные каналы.
Натрий-глюкоза транспортные белки
Белок транспорта натрия-глюкозы использует вторичный активный транспорт для перемещения глюкозы в клетки. Они активны в клетках кишечника и почек, которые должны перемещать глюкозу в системы организма против градиента концентрации.
Эта операция требует энергии, потому что рассматриваемые клетки имеют более высокую концентрацию глюкозы, чем внеклеточной жидкости, Следовательно, глюкоза не сможет самостоятельно диффундировать в клетки; энергия должна быть приложена.
В этом случае энергия поступает из градиента концентрации натрия. Благодаря действию натриево-калиевого насоса, натрий находится вне клетки гораздо больше, чем внутри нее. Таким образом, существует сильный градиент концентрации, способствующий движению натрия в клетку.
Закрытые ионные каналы в улитке
Закрытые ионные каналы пассивный транспорт белки, которые открываются в ответ на конкретные раздражители. Возможно, вы знакомы с ионно-управляемыми ионными каналами, такими как те, которые вызывают срабатывание наших нейронов в ответ на сигналы, поступающие от других нейронов.
Менее известными являются стробированные ионные каналы улитки, которые открываются механическим давлением вместо изменений напряжения. Эти замечательные ионные каналы позволяют нервам нашего внутреннего уха срабатывать в ответ на вибрации звука. Вот как мы слышим.
Внутренние волосковые клетки, с другой стороны, имеют особую работу. В ответ на эти вибрации они открывают ионные каналы в своих клеточных мембранах и освобождают нейротрансмиттеры, как это делает нейрон.
Эти нейротрансмиттеры вызывают укол соседних нервов. И вот как звук преобразуется в нервные импульсы!
- Активный транспорт – Транспорт веществ через биологические мембраны, что требует от клетки расходовать энергию.
- Закрытый ионный канал – Белок, который позволяет ионам проходить, когда открыт, который открывается в ответ на определенный стимул.
- Пассивный транспорт – Транспорт веществ через биологические мембраны, который происходит естественным путем, без необходимости тратить энергию.
викторина
Ответ на вопрос № 1
2. В чем разница между первичным и вторичным активным транспортом?A. Первичный активный транспорт использует белки-носители, тогда как вторичный активный транспорт использует белки-каналы.B. Первичный активный транспорт может транспортировать только одно вещество за раз, тогда как вторичный активный транспорт может транспортировать два.C. Первичный активный транспорт требует энергии; вторичного активного транспорта нет.D. Первичные активные транспортные белки используют АТФ напрямую. Вторичные активные транспортные белки используют энергию, которая получается из других АТФ-зависимых процессов.
Ответ на вопрос № 2
D верно. Все виды активного транспорта требуют от клетки расходовать энергию. Первичные активные транспортные белки берут энергию непосредственно из АТФ; вторичные активные транспортные белки используют энергию от процессов, происходящих из АТФ.
3. Что из перечисленного НЕ является примером активного транспорта?A. Натриево-калиевый насос перемещает натрий и калий как против градиента их концентрации.B. Ионные каналы волосковых клеток открываются в ответ на давление, позволяя ионам проходить через них.C. Транспортер натрия-глюкозы использует градиент концентрации натрия для перемещения глюкозы в клетку.D. Ни один из вышеперечисленных.
Ответ на вопрос № 3
В верно. Ионные каналы являются формой пассивного транспорта; они позволяют ионам двигаться вниз по градиенту концентрации, что не требует затрат энергии.
Читайте также: