
Согласно закону т моргана гены наследуются преимущественно вместе если они расположены в
Обновлено: 02.05.2024
Хромосомная теория наследственности [1] — теория, согласно которой хромосомы, заключённые в ядре клетки, являются носителями генов и представляют собой материальную основу наследственности, то есть преемственность свойств организмов в ряду поколений определяется преемственностью их хромосом. Хромосомная теория наследственности возникла в начале 20 в. на основе клеточной теории и использовалась для изучения наследственных свойств организмов гибридологического анализа.
Содержание
Формирование хромосомной теории
В 1902-1903 гг. американский цитолог У. Сеттон и немецкий цитолог и эмбриолог Т. Бовери независимо друг от друга выявили параллелизм в поведении генов и хромосом в ходе формирования гамет и оплодотворения. Эти наблюдения послужили основой для предположения, что гены расположены в хромосомах. Однако экспериментальное доказательство локализации конкретных генов в конкретных хромосомах было получено только в 1910 г. американским генетиком Т. Морганом, который в последующие годы (1911—1926) обосновал хромосомную теорию наследственности. Согласно этой теории, передача наследственной информации связана с хромосомами, в которых линейно, в определенной последовательности, локализованы гены. Таким образом, именно хромосомы представляют собой материальную основу наследственности.
Формированию хромосомной теории способствовали данные, полученные при изучении генетики пола, когда были установлены различия в наборе хромосом у организмов различных полов.
Генетика пола
Пол, как и любой другой признак организма, наследственно детерминирован. Важнейшая роль в генетической детерминации пола и в поддержании закономерного соотношения полов принадлежит хромосомному аппарату.
У раздельнополых организмов (животных и двудомных растении) соотношение полов обычно составляет 1:1, то есть мужские и женские особи встречаются одинаково часто. Это соотношение совпадает с расщеплением в анализирующем скрещивании, когда одна из скрещиваемых форм является гетерозиготной (Аа), а другая — гомозиготной по рецессивным аллелям (аа). В потомстве в этом случае наблюдается расщепление в отношении 1Аа:1аа. Если пол наследуется по такому же принципу, то вполне логично было бы предположить, что один пол должен быть гомозиготным, а другой — гетерозиготным. Тогда расщепление по полу должно быть в каждом поколении равным 1:1, что и наблюдается в действительности.
При изучении хромосомных наборов самцов и самок ряда животных между ними были обнаружены некоторые различия. Как у мужских, так и у женских особей во всех клетках имеются пары одинаковых (гомологичных) хромосом, но по одной паре хромосом они различаются. Так, у самки дрозофилы имеются две палочковидные хромосомы, а у самца — одна такая же палочковидная, а вторая, парная первой, — изогнутая. Такие хромосомы, по которым самцы и самки отличаются друг от друга, называют половыми хромосомами. Те из них, которые являются парными у одного из полов, называют X-хромосомами (например, у дрозофилы и млекопитающих) или Z-хромосомами (например, у птиц). Непарная половая хромосома, имеющаяся у особей только одного пола, была названа У-хромосомой (у дрозофилы и млекопитающих) или W-хромосомой (у птиц). Хромосомы, в отношении которых между самцами и самками нет различий, называют аутосомами. Следовательно, у дрозофилы особи обоих полов имеют по шесть одинаковых аутосом плюс две половые хромосомы (ХХ у самок и XY у самцов).
Пол, имеющий различные половые хромосомы (X и У), образует гаметы двух типов (половина с X-хромосомой и половина с У-хромосомой), то есть, является гетерогаметным, а пол, содержащий в каждой клетке одинаковые половые хромосомы (X-хромосомы), — гомогаметным.
Открытие половых хромосом и установление их роли в определении пола послужило важным доводом в пользу того, что хромосомы определяют признаки организма.
Определение пола
От чего же зависит рождение мужских и женских особей? Рассмотрим это на примере определения пола у дрозофилы. В ходе гаметогенеза у самок образуется один тип гамет, содержащий гаплоидный набор аутосом и одну X-хромосому. Самцы образуют два типа гамет, половина из которых содержит три аутосомы и одну X-хромосому (ЗА+Х), а половина — три аутосомы и одну У-хромосому (ЗА+У). При оплодотворении яйцеклеток (ЗА+Х) сперматозоидами с X-хромосомами будут формироваться самки (6А+ХХ), а от слияния яйцеклеток со сперматозоидами, несущими У-хромосому, — самцы (6A+XY). Поскольку число мужских гамет с X- и У-хромосомами одинаково, то и количество самцов и самок тоже одинаково. В данном случае пол организма определяется в момент оплодотворения и зависит от хромосомного набора зиготы.
Сходный способ определения пола (XY-тип) присущ всем млекопитающим, в том числе и человеку, клетки которого содержат 44 аутосомы и две X-хромосомы у женщин либо XY-хромосомы у мужчин.
Таким образом, XY-тип определения пола, или тип дрозофилы и человека, — самый распространенный способ определения пола, характерный для большинства позвоночных и некоторых беспозвоночных. Х0-тип встречается у большинства прямокрылых, клопов, жуков, пауков, у которых Y-хромосомы нет вовсе, так что самец имеет генотип Х0, а самка — XX.
У всех птиц, большинства бабочек и некоторых пресмыкающихся самцы являются гомогаметным полом, а самки —- гетерогаметным (типа XY или типа ХО). Половые хромосомы у этих видов обозначают буквами Z и W, чтобы выделить таким образом данный способ определения пола; при этом набор хромосом самцов обозначают символом ZZ, а самки — символом ZW или Z0.
Доказательства того, что половые хромосомы определяют пол организма, были получены при изучении нерасхождения половых хромосом у дрозофилы. Если в одну из гамет попадут обе половые хромосом, а в другую — ни одной, то при слиянии таких гамет с нормальными могут получиться особи с набором половых хромосом ХХХ, ХО, ХХУ и др. Выяснилось, что у дрозофилы особи с набором ХО — самцы, а с набором ХХУ — самки (у человека — наоборот). Особи с набором ХХХ имеют гипертрофированные признаки женского пола (сверхсамки). (Особи со всеми этими хромосомными аберрациями у дрозофилы стерильны). В дальнейшем было доказано, что у дрозофилы пол определяется соотношением (балансом) между числом X-хромосом и числом наборов аутосом.
Наследование признаков, сцепленных с полом
В том случае, когда гены, контролирующие формирование того или иного признака, локализованы в аутосомах, наследование осуществляется независимо от того, кто из родителей (мать или отец) является носителем изучаемого признака. Если же гены находятся в половых хромосомах, характер наследования признаков резко изменяется. Например, у дрозофилы гены, локализованные в X-хромосоме, как правило, не имеют аллелей в У-хромосоме. По этой причине рецессивные гены в X-хромосоме гетерогаметного пола практически всегда проявляются, будучи в единственном числе.
Признаки, гены которых локализованы в половых хромосомах, называются признаками, сцепленными с полом. Явление наследования, сцепленного с полом, было открыто Т. Морганом у дрозофилы.
Х- и У-хромосомы у человека имеют гомологичный (псевдоаутосомный) участок, где локализованы гены, наследование которых не отличается от наследования аутосомных генов.
Помимо гомологичных участков, X- и У-хромосомы имеют негомологичные участки. Негомологичный участок У-хромосомы, кроме генов, определяющих мужской пол, содержит гены перепонок между пальцами ног и волосатых ушей у человека. Патологические признаки, сцепленные с негомологичным участком У-хромосомы, передаются всем сыновьям, поскольку они получают от отца У-хромосому.
Негомологичный участок X-хромосомы содержит в своем составе ряд важных для жизнедеятельности организмов генов. Поскольку у гетерогаметного пола (ХУ) X-хромосома представлена в единственном числе, то признаки, определяемые генами негомологичного участка X-хромосомы, будут проявляться даже в том случае, если они рецессивны. Такое состояние генов называется гемизиготным. Примером такого рода X-сцепленных рецессивных признаков у человека являются гемофилия, мышечная дистрофия Дюшена, атрофия зрительного нерва, дальтонизм (цветовая слепота) и др.
Гемофилия — это наследственная болезнь, при которой кровь теряет способность свертываться. Ранение, даже царапина или ушиб, могут вызвать обильные наружные или внутренние кровотечения, которые нередко заканчиваются смертью. Это заболевание встречается, за редким исключением, только у мужчин. Было установлено, что обе наиболее распространенные формы гемофилии (гемофилия А и гемофилия В) обусловлена рецессивными генами, локализованными в X-хромосоме. Гетерозиготные по данным генам женщины (носительницы) обладают нормальной или несколько пониженной свертываемостью крови.
Фенотипическое проявление гемофилии у девочек будет наблюдаться в том случае, если мать девочки является носительницей гена гемофилии, а отец — гемофиликом. Подобная закономерность наследования характерна и для других рецессивных, сцепленных с полом признаков.
Сцепленное наследование
Независимое комбинирование признаков (третий закон Менделя) осуществляется при условии, что гены, определяющие эти признаки, находятся в разных парах гомологичных хромосом. Следовательно, у каждого организма число генов, способных независимо комбинироваться в мейозе, ограничено числом хромосом. Однако в организме число генов значительно превышает количество хромосом. Например, у кукурузы до эры молекулярной биологии было изучено более 500 генов, у мухи дрозофилы — более 1 тыс., а у человека — около 2 тыс. генов, тогда как хромосом у них 10, 4 и 23 пары соответственно. То, что число генов у высших организмов составляет несколько тысяч, было ясно уже У. Сэттону в начале XX века. Это дало основание предположить, что в каждой хромосоме локализовано множество генов. Гены, локализованные в одной хромосоме, образуют группу сцепления и наследуются вместе.
Совместное наследование генов Т. Морган предложил назвать сцепленным наследованием. Число групп сцепления соответствует гаплоидному числу хромосом, поскольку группу сцепления составляют две гомологичные хромосомы, в которых локализованы одинаковые гены. (У особей гетерогаметного пола, например, у самцов млекопитающих, групп сцепления на самом деле на одну больше, так как X- и У-хромосомы содержат разные гены и представляют собой две разные группы сцепления. Таким образом, у женщин 23 группы сцепления, а у мужчин — 24).
Способ наследования сцепленных генов отличается от наследования генов, локализованных в разных парах гомологичных хромосом. Так, если при независимом комбинировании дигетерозиготная особь образует четыре типа гамет (АВ, Ab, аВ и ab) в равных количествах, то при сцепленном наследовании (в отсутствие кроссинговера) такая же дигетерозигота образует только два типа гамет: (АВ и ab) тоже в равных количествах. Последние повторяют комбинацию генов в хромосоме родителя.
Было установлено, однако, что кроме обычных (некроссоверных) гамет возникают и другие (кроссоверные) гаметы с новыми комбинациями генов — Ab и аВ, отличающимися от комбинаций генов в хромосомах родителя. Причиной возникновения таких гамет является обмен участками гомологичных хромосом, или кроссинговер.
Кроссинговер происходит в профазе I мейоза во время конъюгации гомологичных хромосом. В это время части двух хромосом могут перекрещиваться и обмениваться своими участками. В результате возникают качественно новые хромосомы, содержащие участки (гены) как материнских, так и отцовских хромосом. Особи, которые получаются из таких гамет с новым сочетанием аллелей, получили название кроссинговерных или рекомбинантных.
Частота (процент) перекреста между двумя генами, расположенными в одной хромосоме, пропорциональна расстоянию между ними. Кроссинговер между двумя генами происходит тем реже, чем ближе друг к другу они расположены. По мере увеличения расстояния между генами все более возрастает вероятность того, что кроссинговер разведет их по двум разным гомологичным хромосомам.
Расстояние между генами характеризует силу их сцепления. Имеются гены с высоким процентом сцепления и такие, где сцепление почти не обнаруживается. Однако при сцепленном наследовании максимальная частота кроссинговера не превышает 50 %. Если же она выше, то наблюдается свободное комбинирование между парами аллелей, не отличимое от независимого наследования.
Биологическое значение кроссинговера чрезвычайно велико, поскольку генетическая рекомбинация позволяет создавать новые, ранее не существовавшие комбинации генов и тем самым повышать наследственную изменчивость, которая дает широкие возможности адаптации организма в различных условиях среды. Человек специально проводит гибридизацию с целью получения необходимых вариантов комбинаций для использования в селекционной работе.
Понятие о генетической карте
Т. Морган и его сотрудники К. Бриджес, А. Г. Стертевант и Г. Дж. Меллер экспериментально показали, что знание явлений сцепления и кроссинговера позволяет не только установить группу сцепления генов, но и построить генетические карты хромосом, на которых указаны порядок расположения генов в хромосоме и относительные расстояния между ними.
Генетической картой хромосом называют схему взаимного расположения генов, находящихся в одной группе сцепления. Такие карты составляются для каждой пары гомологичных хромосом.
Возможность подобного картирования основана на постоянстве процента кроссинговера между определенными генами. Генетические карты хромосом составлены для многих видов организмов: насекомых (дрозофила, комар, таракан и др.), грибов (дрожжи, аспергилл), для бактерий и вирусов.
Наличие генетической карты свидетельствует о высокой степени изученности того или иного вида организма и представляет большой научный интерес. Такой организм является прекрасным объектом для проведения дальнейших экспериментальных работ, имеющих не только научное, но и практическое значение. В частности, знание генетических карт позволяет планировать работы по получению организмов с определенными сочетаниями признаков, что теперь широко используется в селекционной практике. Так, создание штаммов микроорганизмов, способных синтезировать необходимые для фармакологии и сельского хозяйства белки, гормоны и другие сложные органические вещества, возможно только на основе методов генной инженерии, которые, в свою очередь, базируются на знании генетических карт соответствующих микроорганизмов.
Генетические карты человека также могут оказаться полезными в здравоохранении и медицине. Знания о локализации гена в определенной хромосоме используются при диагностике ряда тяжелых наследственных заболеваний человека. Уже теперь появилась возможность для генной терапии, то есть для исправления структуры или функции генов.
Сравнение генетических карт разных видов живых организмов способствует также пониманию эволюционного процесса.
Основные положения хромосомной теории наследственности
Анализ явлений сцепленного наследования, кроссинговера, сравнение генетической и цитологической карт позволяют сформулировать основные положения хромосомной теории наследственности:


Сцепленное наследование. Г. Мендель опубликовал результаты своих исследований в 1865 г., однако тогда его открытия остались незамеченными. Только в 1900 г. К-Корренс (Германия), Г. де Фриз (Голландия) и Э. Чер мак (Австрия) независимо друг от друга обнаружили у разных видов растений те же закономерности наследования признаков, что и Г. Мендель. Английский генетик У. Бэтсон подтвердил законы Менделя на животных. Переоткрытие законов Менделя вызвало глубокий интерес к изучению закономерностей наследования признаков и способствовало быстрому развитию генетики.
В 1902 г. немецкий цитолог и эмбриолог Т. Б о в е р и представил доказательства участия хромосом в процессах передачи наследственной информации. Он показал, например, что нормальное развитие морского ежа возможно лишь при наличии всех хромосом. Подобную связь заметил в 1903 г. и американский цитологУ. С эттон. Так получили обоснование предположения Менделя
о наследственных факторах, о наличии одинарного набора этих факторов в гаметах и двойного — в зиготах. В 1909 г. датский биолог В. Иогансен ввел понятие s.ген:/.
В 1910 г. американский генетик Т. Морган экспериментально доказал, что гены расположены в хромосомах. Многочисленные исследования Моргана и его учеников привели к целому ряду важнейших открытий, которые легли в основу хромосомной теории наследственности. Одно из ее положений можно сформулировать следующим образом: гены расположены в хромосомах в линейном порядке и занимают определенные участки — локусы, причем аллельные гены находятся в одинаковых локусах гомологичных хромосом.
Закон независимого наследования (третий закон Менделя) справедлив в том случае, если неаллельные гены находятся в разных парах хромосом. Однако количество генов у живых организмов значительно больше числа хромосом. Например, у человека около 25 тыс. генов, а количество хромосом —
23 пары (2п = 46); у плодовой мушки дрозофилы приблизительно 14 тыс. генов и всего 4 пары хромосом (2п = 8). Следовательно, каждая хромосома содержит множество генов. Будут ли гены, локализованные в одной хромосоме, наследоваться независимо? Очевидно, что нет.
Гены, расположенные в одной хромосоме, образуют группу сцепления и наследуются вместе. Совместное наследование генов Т. Морган предложил называть сцепленным наследованием (в отличие от независимого). Каждая пара гомологичных хромосом содержит гены, контролирующие одни и те же признаки, поэтому количество групп сцепления равно числу пар хромосом. Например, у человека 23 группы сцепления, а у дрозофилы — 4.

Вам известно, что при независимом наследовании дигетерозиготная особь, например =^=, образует четыре типа гамет в равном соотношении, т. е. по 25 %: Л В, АЬ, а В и ab. Это обусловлено тем, что неаллельные гены находятся в разных парах хромосом. Если же они расположены в гомологичных хромосомах, следовало бы ожидать, что дигетерозигота будет производить лишь два типа га-
мет: 50 % АВ и 50 % ab (обратите внимание на то, что сцепленные гены записываются в одну хромосому).
Однако Т. Морган обнаружил, что в большинстве случаев дигетерозиготные особи образуют не два, а четыре типа гамет. Помимо ожидаемых АВ и ab формируются также гаметы с новыми комбинациями генов: АЬ и аВ, только в меньшем процентном соотношении. Рассмотрим один из экспериментов Т. Моргана, в котором изучалось наследование сцепленных генов у дрозофилы.

Если почистить фрукты или овощи и не сразу выбросить очистки либо оставить фрукты на столе на несколько дней, то можно заметить, как вокруг остатков пищи начнут роиться маленькие мушки размером около 2—3,5 мм. Это дрозофилы — плодовые мушки, род насекомых отряда Двукрылые (рис. 95). Обычно дрозофилы имеют красные глаза и желтокоричневую окраску брюшка. Жизненный цикл дрозофил короток: развитие от яйца до половозрелой особи при 25 °С занимает 10 дней. Небольшие размеры, высокая плодовитость, простота культивирования и ряд других особенностей на долгое время сделали дрозофилу главным объектом генетики. Не один нобелевский лауреат, кроме своего интеллекта, обязан ей своими научными достижениями.
Путем скрещивания чистой линии дрозофил, имеющих серое тело и нормальные (длинные) крылья, с чистой линией, особи которой имели черное тело и зачаточные крылья, были получены гибриды первого поколения (рис. 96). Все они в соответствии с законом единообразия были серыми с нормально развитыми крыльями. Следовательно, у дрозофил серое тело (А) полностью доминирует над черным (а), а нормальные крылья (В) — над зачаточными (b). Все гибриды первого поколения — дигетерозиготы.
Затем было проведено анализирующее скрещивание (рис. 97). Дигетерозиготную самку из гибридного поколения скрестили с рецессивным дигомозиготным самцом (черное тело и зачаточные крылья). В потомстве было получено по 41,5 % особей с серым телом, нормальными крыльями и черным телом, зачаточными крыльями, а также по 8,5 % мух с серым телом, зачаточными крыльями и черным телом, нормальными крыльями.

Если бы гены, определяющие цвет тела и развитие крыльев, находились в разных парах хромосом, соотношение фенотипических классов было бы равным — по 25 %. Но этого не наблюдалось, значит, гены находятся в гомологичных хромосомах и наследуются сцепленно.

Несмотря на сцепление генов, АВ самка производила не два, а четыре типа гамет. Однако гамет с исходными сочетаниями сцепленных генов формировалось намного больше (АВ и ab вместе составили 83 %), чем с новыми их сочетаниями (сумма АЬ и дВ равна 17 %).
Было выяснено, что причиной появления хромосом с новыми комбинациями родительских генов является кроссинговер. Вы помните, что этот процесс происходит в профазе I мейоза и представляет собой обмен соответствующими участками между гомологичными хромосомами. Таким образом, кроссинговер препятствует полному (абсолютному) сцеплению генов. Гаметы, которые образуются в результате кроссинговера, и особи, которые развиваются при участии таких гамет, называются кроссоверными или рекомбинантными. В рассмотренном эксперименте гаметы АЬ и аВ являлись кроссоверными, а гаметы АВ и ab — некроссоверными (см. рис. 97).
Кроссинговер между конкретными сцепленными генами происходит с определенной вероятностью (частотой). Для расчета частоты кроссинговера (rf, от англ. recombination frequency — частота рекомбинации) можно пользоваться следующей формулой:
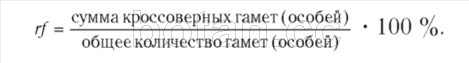
Таким образом, между генами А и В, контролирующими цвет тела и длину крыльев дрозофилы, кроссинговер происходит с частотой: rfAB= 17 %.
Дальнейшие исследования, проведенные Т. Морганом и его сотрудниками, показали, что частота кроссинговера пропорциональна расстоянию между генами, расположенными в одной хромосоме. Чем больше расстояние между сцепленными генами, тем чаще между ними происходит кроссинговер. И наоборот, чем ближе друг к другу расположены гены, тем меньше частота кроссинговера между ними. Чем объясняется эта закономерность?
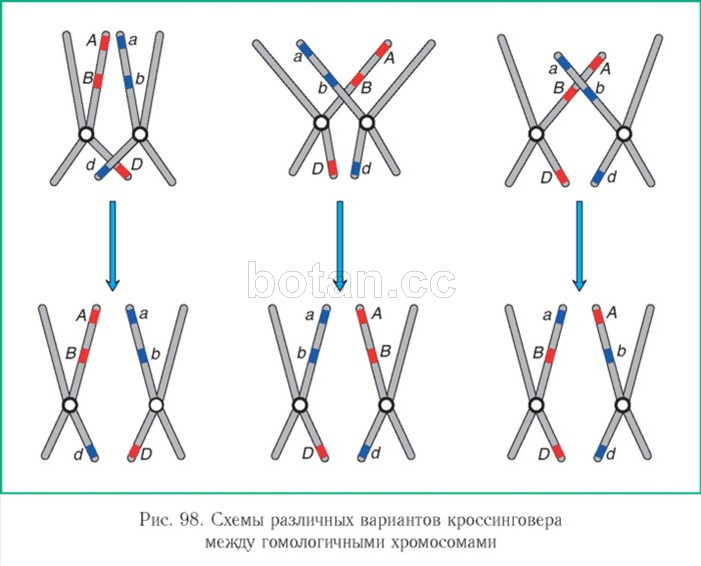
В профазе I мейоза при конъюгации гомологичных хромосом образование перекрестов между хроматидами осуществляется произвольно, на любых соответствующих участках. Рассмотрим рисунок 98.
Гены А и В (или а. и Ь) находятся сравнительно близко друг к другу. Вероятность того, что перекрест произойдет именно на участке, разделяющем эти гены, невелика. Гены А и D (или а. и d) располагаются на значительном расстоянии друг от друга. Поэтому вероятность того, что хроматиды перекрестятся на каком-либо участке между ними, намного выше. Значит, чем больше расстояние между генами, тем чаще они разделяются при кроссинговере.
Таким образом, частота кроссинговера позволяет судить о расстоянии между генами. В честь Т. Моргана единица измерения расстояния между генами получила название моргай и да или, что то же самое, санти моргай и да (сМ).
Морганида (сантиморганида, сМ) — это генетическое расстояние, на котором кроссинговер происходит с вероятностью 1 %.
Биологическое значение кроссинговера чрезвычайно велико. В результате этого процесса возникают новые комбинации родительских генов, что повышает генетическое разнообразие потомства и расширяет возможности адаптации организмов к различным условиям окружающей среды.
Генетические карты. Т. Морган и сотрудники его лаборатории показали, что знание частоты кроссинговера между сцепленными генами позволяет строить генетические карты хромосом. Генетическая карта представляет собой схему взаимного расположения генов, находящихся в одной группе сцепления, с учетом расстояний между ними (рис. 99).
Генетические карты хромосом уже составлены для человека, многих видов животных, растений, грибов и микроорганизмов. Наличие генетической карты свидетельствует о высокой степени изученности того или иного вида организма и представляет большой научный интерес. Такой организм является прекрасным объектом для проведения дальнейших экспериментальных работ, имеющих не только научное, но и практическое значение. В частности, знание генетических карт позволяет планировать работы по получению организмов с определенными сочетаниями признаков, что широко применяется в селекционной практике. Генетические карты хромосом человека используются в медицине для диагностики и лечения ряда наследственных заболеваний.

Основные положения хромосомной теории наследственности.
1. Гены в хромосомах расположены линейно, в определенной последовательности. Аллельные гены находятся в одинаковых локусах гомологичных хромосом.
2. Гены, расположенные в одной хромосоме, образуют группу сцепления и наследуются вместе. Количество групп сцепления равно числу пар хромосом.
3. Сцепление генов может нарушаться в результате кроссинговера, происходящего при конъюгации гомологичных хромосом в профазе I мейоза.
4. Частота кроссинговера пропорциональна расстоянию между генами: чем больше расстояние, тем выше частота кроссинговера, и наоборот.
Б. За единицу расстояния между сцепленными генами принята 1 морганида — расстояние, на котором кроссинговер происходит с вероятностью 1 %.
1. Всегда ли соблюдается III закон Г. Менделя? В каких случаях гены наследуются независимо, а в каких случаях — сцепленно?
2. Что такое группа сцепления? Чему равно количество групп сцепления в клетках разных организмов?
3. Почему частота кроссинговера между сцепленными генами зависит от расстояния между ними?
4. Что такое генетические карты хромосом? Каковы перспективы их использования?
5. Сформулируйте основные положения хромосомной теории наследственности.
6. Какие типы гамет и в каком процентном соотношении будут формировать дигетерозиготные особи - и ——, если известно, что расстояние между генами А и В составляет 20 морган ид?
7. Серый цвет тела у дрозофилы доминирует над желтым, красные глаза — над гранатовыми. Гены, отвечающие за эти признаки, локализованы в первой паре хромосом и находятся на расстоянии 44 морганиды. Скрестили чистые линии серотелых мух с гранатовыми глазами и желтотелых с красными глазами. Из полученных гибридов выбрали самку и подвергли ее анализирующему скрещиванию. Каким будет процентное соотношение фенотипических классов в потомстве?
8. У одного из видов растений рассеченные листья доминируют над цельными, а синяя окраска цветков — над розовой. В результате анализирующего скрещивания было получено потомство четырех фенотипических классов: 1) 133 растения с рассеченными листьями и синими цветками; 2) 362 растения с рассеченными листьями и розовыми цветками; 3) 127 растений с цельными листьями и розовыми цветками; 4) 378 растений с цельными листьями и синими цветками. Затем растения первого фенотипического класса скрестили с растениями второго фенотипического класса. Сколько процентов полученных гибридов будут иметь рассеченные листья и розовые цветки? Цельные листья и синие цветки?
Глава 1. Химические компоненты живых организмов
Глава 2. Клетка — структурная и функциональная единица живых организмов
Глава 3. Обмен веществ и преобразование энергии в организме
Глава 4. Структурная организация и регуляция функций в живых организмах
Глава 5. Размножение и индивидуальное развитие организмов
Глава 6. Наследственность и изменчивость организмов
Глава 7. Селекция и биотехнология
Биология: учеб. для 10-го кл. учреждений общ. сред, образования с рус. яз. обуч. / Н. Д. Лисов [и др.]; под ред. Н. Д. Лисова. — 3-е изд., перераб. — Минск : Народная асвета, 2014. — 270 с.: ил.
Разработки уроков на конкурс “Учитель - учителю”. Уроки 7, 8.
Явление сцепленного наследования
Закон Томаса Моргана
- раскрыть сущность сцепленного наследования;
- сформировать знания о группах сцепления и генетических картах;
- объяснить причину фенотипического расщепления 3:1;
- выявить причину появления рекомбинантных фенотипов;
- освоить решение задач по кроссинговеру.
Форма урока – лекция.
Понятия урока: сцепленное наследование, группы сцепления, генетические карты хромосом, рекомбинация генов.
- Индивидуальный опрос (работа у доски):
- решить задачи с комментариями № 6,7 с. 105 учебника.
- Фронтальный опрос
- беседа по вопросам 1, 3 параграфа 25.
Домашнее задание: §26, задача №2 с. 107.
II. Изучение нового материала
- Группы сцепления.
- Работа Томаса Моргана.
- Решение задач.
Все примеры, которые мы рассматривали на предыдущих уроках, относились к наследованию генов, находящихся в разных хромосомах. Однако любой организм имеет тысячи признаков, а количество хромосом (в которых находятся гены, определяющие эти признаки), ограничено. Таким образом, в каждой хромосоме должно находиться большое количество генов.
Соматические клетки человека содержат 46 хромосом, таким образом мы имеем ситуацию, когда относительно небольшое количество хромосом содержат большое разнообразие признаков, определяемых генами. Т.е. определенные признаки (гены) группируются в определенных хромосомах вместе. В генетике такие группы генов, лежащие в одной хромосоме, называют сцепленными .
Все гены какой-либо одной хромосомы образуют группу сцепления ; они обычно наследуются вместе, т.к. попадают в одну гамету. Гены, образующие группу сцепления, не подчиняются закону Г. Менделя о независимом распределении. Тогда как же Г. Мендель мог сделать свои открытия? На лицо случай научного везения – он случайно выбрал признаки, которые определяются генами разных хромосом.

Цвет семян (хромосома) Форма семян (хромосома)
А мы сегодня рассматриваем другой случай:

Работа Томаса Моргана
Большую работу по изучению наследования сцепленных генов провел американский генетик Томас Морган.
Итак, у мухи гены представлены следующими парами аллелей:
Однако Томас Морган получил другое соотношение фенотипов – 3 : 1. Почему это произошло?
Генетическое объяснение отношения фенотипов 3 : 1

Скрещивание потолков F 1


На самом деле, в результате скрещивания мух с исходной комбинацией признаков (серое тело – длинные крылья, темное тело – зачаточные крылья) оказывается значительно больше, чем мух с перекомбинированными признаками. Томас Морган получал следующее соотношение:
Серые с длинными крыльями – 41.5 %
Черные с зачаточными крыльями – 41.5 %
Серые с зачаточными крыльями – 8.5 %
Черные с длинными крыльями – 8.5 %
О чем это говорит?
Гены, обусловившие признаки серое тело – длинные крылья, темное тело – зачаточные крылья, наследуются преимущественно вместе, или, иначе говоря, оказываются сцепленными между собой.
К каким же выводам мы пришли?
- Полное скрещивание встречается редко, образуются особи с совершенно новыми признаками – их новые фенотипы называются рекомбинантными (они образовались во время мейоза при кроссинговере ).
- Оба гена (длины и цвета) находятся в 1 хромосоме.
Вспомним, что называется кроссинговер? От чего зависит частота кроссинговера? Решим задачу:

A, B, C – три генных локуса, расположенных в одной хромосоме. Вероятность кроссинговера для генов A и C выше, чем для A и B, т.к. частота кроссинговера зависит от расстояния между генами.

Изображены локусы, 12 аллелей, расположенных в паре хромосомы. Показаны их расстояния от центромер.
а) Как называются эти хромосомы? (гомологичными)
б) Между какими локусами чаще будет происходить кроссинговер? (1 и 6)
в) произойдет ли кроссинговер между генами, определяющими цвет глаз и форму усиков дрозофил? (на этот вопрос нельзя ответить, т.к. не указан локус гена, опредляющего форму усиков).
Работы Т. Моргана позволяют составить схемы или карту расположения генов. Такая генетическая карта показывает последовательность размещения генов в хромосоме и расстояния между ними. В генетике принято расстояние между генами, расположенными в одной хромосоме, определять в процентах гамет, при образовании которых в результата кроссинговера произошла перекомбинация генов в гомологичных хромосомах. За единицу расстояния между генами, находящимися в одной хромосоме, принят 1% кроссинговера. Такая единица в честь Т. Моргана, получила название морганиды . Если полученные таким способом расстояния между генами нанести на прямую, то возникнет схема расположения генов на хромосоме – хромосомная карта (генетическая карта).
Сертификат и скидка на обучение каждому участнику

УЧЕБНАЯ ДИСЦИПЛИНА: БИОЛОГИЯ
Тема: Хромосомная теория наследственности
Ознакомиться с лекционным материалом по теме
Задача 1. От скрещивания между гомозиготным серым длиннокрылым самцом дрозофилы и гомозиготной черной самкой с зачаточными крыльями в F1 получено потомство с серым телом и длинными крыльями. Гены окраски тела и длины крыльев наследуются сцеплено и расстояние между ними равно 19 морганид (M). Какова вероятность в % появления черных мух с длинными крыльями при скрещивании самки дрозофилы из поколения F1 с черным самцом, имеющим зачаточные крылья.
Задача 2. У томатов ген A определяет высокий рост стебля, а ген a – карликовость. Ген B детерминирует шаровидную форму плодов, а ген b – грушевидную их форму. Гены высоты стебля и формы плодов сцеплены и находятся на расстоянии 20 M. Скрестили гетерозиготное по обоим признакам растение с карликовым растением, имеющем грушевидные плоды. Какие генотипы, и в каком процентном соотношении можно получить в результате данного скрещивания?
Задача 3 . Резус-положительность и аномальная форма эритроцитов определяются доминантными аутосомными генами, соответственно, Rh + и A, расположенными на расстоянии 3-х морганид (М) друг от друга. Один из супругов гетерозиготен по обоим признакам. При этом резус-положительность он унаследовал от одного родителя, а аномальную форму эритроцитов – от другого. Второй супруг резус-отрицателен (rh - ) и имеет нормальные эритроциты (a). Какова вероятность в процентах рождения в семье ребенка, имеющего положительный резус-фактор и эритроциты нормальной формы?
Сцепленное наследование генов. Хромосомная теория наследственности Т. Моргана
Согласно Г. Менделю гены наследуются независимо и располагаются в разных хромосомах. Дигетерозиготу AaBb в виде цитологической формулы можно представить следующим образом:
В ходе мейоза она формирует 4 типа гамет: AB, Ab, aB, ab.
В 1911–1912 гг. американский генетик Т. Морган доказал, что гены, расположенные в одной хромосоме, могут наследоваться сцеплено (вместе). Это обусловлено тем, что генов намного больше, чем хромосом. Совместное наследование генов, ограничивающее их свободное комбинирование, Т. Морган назвал сцеплением генов, или сцепленным наследованием. В случае, когда гены наследуются сцепленно, генотип дигетерозиготы AaBb с помощью цитологической формулы можно представить следующим образом:
В ходе мейоза, если имеет место полное сцепление генов, она формирует 2 типа гамет – AB, ab. Такие гаметы называются некроссоверными.
Исследования Т. Моргана и его учеников показали, что в гомологичной паре хромосом регулярно происходит обмен генами. Процесс обмена идентичными участками гомологичных хромосом с находящимися в них генами называют перекрестом хромосом, или кроссинговером. Кроссинговер обеспечивает новые сочетания генов, расположенных в гомологичных хромосомах. Поэтому, благодаря явлению кроссинговера, помимо некроссоверных, формируются также кроссоверные гаметы – Ab, аВ.
Закон Т. Моргана формулируется следующим образом: гены, локализованные в одной хромосоме, наследуются сцеплено (вместе), причем сила сцепления зависит от расстояния между генами.
Основные положения хромосомной теории Т. Моргана:
1) гены локализованы в хромосомах, и различные хромосомы содержат неодинаковое число генов. Набор генов в каждой из негомологичных хромосом уникален;
- гены расположены в хромосоме в определенной линейной последовательности;
- гены образуют группы сцепления и могут наследоваться сцепленно. Число групп сцепления соответствует гаплоидному набору хромосом (у человека 23 группы сцепления);
4) между гомологичными хромосомами возможен кроссинговер (перекрест) и обмен аллельными генами, причем перекрест происходит тогда, когда есть расстояние между генами;
5) расстояние между генами в хромосоме пропорционально частоте перекреста и выражается в процентах кроссинговера.
Процент кроссинговера между генами рассчитывается по формуле:
a– число кроссоверных особей одного класса; b – число кроссоверных особей другого класса;
n – общее число особей; Х – % кроссинговера между генами (расстояние между генами в % кроссинговера).
Расстояние между генами измеряется в специальных единицах – морганидах. 1 морганида (1 M) = 1 % кроссинговера.
В генетических исследованиях использовали мушку-дрозофилу, имеющую кариотип, состоящий из 8 хромосом, обладающую плодовитостью, множеством альтернативных признаков. Т. Морган рассмотрел ситуацию, когда полное сцепление генов и нарушение сцепления генов (кроссинговер) происходят у особи одновременно. На первом этапе для скрещивания взяты гомозиготные особи (P ♀AABB x ♂aabb). В результате в первом поколении все особи оказались дигетерозиготными (AaBb).
Ген Признак
A серая окраска тела
а темная окраска тела
В длинные крылья
b зачаточные крылья.
Полное сцепление генов
G AB, ab ab – гаметы самца
(Некроссоверные гаметы самки)
При полном сцеплении генов наблюдается расщепление 1:1. Всего получено 83 % особей, которые унаследовали фенотип родителей, т. к сформировались из некроссоверных гамет. Среди них особи с серым телом, нормальными крыльями составили 41,5 % и особи с черным телом, зачаточными крыльями также составили 41,5 %.
Нарушение сцепления генов
G Ab, aB ab – гаметы самца
(Кроссоверные гаметы самки)
При неполном сцеплении генов также наблюдается расщепление 1:1. Всего получено 17 % особей с рекомбинантными фенотипами, т. к. они сформировались из кроссоверных гамет. Особи с новым сочетанием признаков называются рекомбинантами. Среди рекомбинантов мухи с серым телом, зачаточными крыльями составили 8,5 %, а с черным телом, нормальными крыльями также составили 8,5 %.
Данные результаты свидетельствуют о наличии сцепления генов и кроссинговера между ними. При сцепленном наследовании генов, протекающем с кроссинговером, во втором поколении количество родительских потомков – 83 % (особей с родительскими фенотипами), а количество рекомбинантных потомков невелико и составляет 17 % . Следовательно, расстояние между генами равно 17 %, или 17 морганидам.
Следует отметить, что негомологичные хромосомы содержат разные группы сцепления, и не конъюгируют при мейозе.
Генетика пола. Теория хромосомного определения пола. Наследование, сцепленное с полом
Пол – совокупность морфологических, биохимических, генетических и физиологических признаков организма и наследуется как любой другой признак. Половые хромосомы – хромосомы, определяющие различие кариотипов у особей разных полов. Кариотипы мужского и женского организмов отличаются по половым хромосомам. Кариотип – диплоидный набор хромосом соматических клеток организма. Кариотип мужского организма включает 22 пары – 44 A (аутосомы) и две половые хромосомы (х, у) и составляет 46 хромосом (рис. 1).
Кариотип женского организма включает 22 пары – 44 A (аутосомы) и две половые хромосомы (Х, Х) и составляет 46 хромосом. Аутосомы у обоих полов аналогичны. В мужском организме 23-я пара не схожа внешне, – одна хромосома меньше, чем другая. У женщин половые хромосомы одного размера.

Рис . 1. Кариотип человека: 22 пары аутосом и половые хромосомы.
Хромосомы расположены в порядке уменьшения их длины и подразделены на 7 групп, обозначенных латинскими буквами A, B, C, D, E, F, G. Каждая пара хромосом имеет порядковый номер (от 1 до 22). Половые хромосомы Х и У выделены отдельно (рис. 1).
Различают гомогаметный (женский) пол и гетерогаметный (мужской) пол.
Гомогаметный пол – это пол, имеющий в каждой соматической клетке, кроме аутосом, две одинаковые половые хромосомы.
Гетерогаметный пол – это пол, который содержит в каждой соматической клетке, кроме аутосом, две разные половые хромосомы.
У гомогаметного пола образуется один тип гамет, а у гетерогаметного– два типа гамет.
♂ 44 A + XY х ♀ 44 A + ХХ
G 22 A + X 22 A + Х
22 A + Y 22 A + Х
Выделяют гетерогаметный пол Х0 и ХY. Если гетерогаметный пол имеет одну Х-хромосому, то это тип X0, а если он имеет пару различающихся половых хромосом – Х и Y, то это тип ХY. Тип ХY – у человека, млекопитающих, дрозофилы, а тип X0 у клопов, кузнечиков. У самок в результате мейоза образуются гаметы, содержащие только Х-хромосому, а у самцов – одни гаметы с X-, а другие с Y-хромосомой. При нарушении мейоза возможно образование гамет без половых хромосом и гамет, содержащих Х и Y хромосомы. Оплодотворение яйцеклетки сперматозоидом, несущим Х-хромосому, приводит к образованию ХХ-зиготы, из которой развивается женская особь. Оплодотворение яйцеклетки сперматозоидом, несущим Y-хромосому, приводит к образованию ХY-зиготы, из которой развивается мужская особь.
У птиц, бабочек, рептилий и амфибий гомогаметным является мужской пол.
Типы хромосомного определения пола
Выделяют следующие типы хромосомного определения пола: ХY – у человека, дрозофилы; Х0 – у клопов, кузнечиков; ZW – у птиц, бабочек, рептилий и амфибий; по плоидности – у пчел, насекомых, тлей.
Схема хромосомного определения пола у человека (тип ХУ)
P ♀ 44 A + ХХ x ♂ 44 A + XY
G 22 A + Х 22 A + Х
22 A + Х 22 A + Y
F1 44 A + ХХ ; 44 A + XY; 44 A + ХХ ; 44 A + XY
Пол человека определяется в момент или после оплодотворения. Из зиготы 44 A + ХХ разовьется организм женского пола, а из зиготы 44 А + XY – организм мужского пола. Соотношение полов составляет 1:1.
Схема хромосомного определения пола у дрозофилы (тип XY)
P ♀ 6 A + XX x ♂ 6 A + Х Y
G 3 A + Х 3 A + Х
F1 6A + ХХ ; 6A + ХХ ; 6A + Х Y; 6A + Х Y
Схема хромосомного определения пола у клопов из рода Protenor (тип Х0)
P♀ 12 A + ХХ x ♂12 A + Х 0
G 6 A + Х ; 6 A + Х ;
F1 12 A + ХХ ; 12 A + Х 0; 12 A + ХХ ; 12 A + Х 0
Схема хромосомного определения пола у птиц (тип ZW)
А ) у кур : P ♀ 76 A + ZW x ♂ 76 A + ZZ
G 38 A + Z; 38 A + Z;
38 A + W 38 A + Z
F1 76 A + ZZ; 76 A + ZZ; 76 A + ZW; 76 A + ZW
Соотношение полов у птиц составляет 1:1
Схема хромосомного определения пола у пчел (по плоидности)
У пчел самцы – гаплоидные (моноплоидные), а самки – диплоидные.
P♂ 16 A + Х x ♀32 A + ХХ
G 16 A + Х ; 16 A + Х ; 16 A + Х
F1 32 A + ХХ; 16 A + Х
Кариотип самки – 32 A + ХХ, а кариотип самца – 16 A + Х. Соотношение полов – 1:1. Особь с кариотипом 16 A +Х развивается партеногенетически (без оплодотворения).
Наследование, сцепленное с полом
Выше было показано, если гены, ответственные за формирование признака, расположены в аутосомах, то наследование осуществляется независимо от того, кто из родителей (мать или отец) является носителем данного признака. Однако признаки могут определяться генами, лежащими в половых хромосомах. В этом случае наследование признака зависит от того, кто из родителей является носителем данного признака.
Ген, расположенный в Х-хромосоме, является сцепленным с полом. Некоторые заболевания передаются по женской линии, через Х-хромосому (гемофилия, дальтонизм и др.).
Гемофилия (несвертываемость крови) определяется рецессивным геном h, а нормальная свертываемость крови детерминируется доминантным геном H. Рассмотрим, какое потомство может появиться у женщины, вступающей в брак со здоровым по этому признаку мужчиной.
Схема наследования гемофилии
P ♀ Х H Х h x ♂ Х H Y
F1 Х H Х H ; Х H Х h ; Х H Y; Х h Y
Вероятность рождения ребенка с гемофилией составляет 25 %. Одна из дочерей является гетерозиготным носителем гена h.
Схема наследования дальтонизма
Дальтонизм (нарушение цветного зрения) определяется рецессивным геном d, а нормальное зрение детерминируется доминантным геном D.
P♀ Х D Х d x ♂ Х D Y
F1 Х D Х D ; Х D Х d ; Х D Y; Х d Y
Вероятность рождения ребенка с дальтонизмом составляет 25 %. Одна из дочерей является гетерозиготным носителем гена d.
Наследование белоглазости у дрозофилы
Доминантный ген W определяет красноглазость у дрозофилы, а рецессивный ген w – белоглазость.
Скрестили гомозиготную белоглазую самку с красноглазым самцом:
P ♀ Х w Х w x ♂Х W Y
F1 Х W Х w ; Х W Х w ; Х w Y; Х w Y
Данная схема скрещивания показывает, что при сцепленном с полом наследовании, дочери наследуют признаки от отцов, а сыновья – от матерей.
Взаимодействие неаллельных генов
Неаллельные гены – гены, расположенные в разных парах гомологичных хромосом (A и b, b и C, A и C, А и B и др.).
Типы взаимодействия неаллельных генов: комплементарность, эпистаз, полимерия.
Комплементарность – такое взаимодействие неаллельных генов, когда два доминантных гена при совместном действии, обусловливают развитие нового фенотипического признака
Пример 1. Наследование пурпурной окраски цветков у душистого горошка
Скрещивали растения горошка с белыми цветками, имеющими генотипы aaBB и AAbb:
P ♀ aaBB x ♂ AAbb
В F1 получены особи с пурпурными цветками. Пурпурная окраска цветков у особей в F1 с генотипом AaBb обусловлена взаимодействием двух доминантных комплементарных генов – A и B.
Читайте также: