
Сколько типов гамет образуется у дигетерозиготной самки при сцепленном наследовании и кроссинговере
Обновлено: 30.06.2024
Сцепление генов – это совместное наследование генов, расположенных в одной и той же хромосоме. Количество групп сцепления соответствует гаплоидному числу хромосом, то есть у дрозофилы 4. Природу сцепленного наследования объяснил Морган с сотрудниками. В качестве объекта исследования они избрали плодовую муху дрозофилу, которая оказалась очень удобной моделью для изучения данного феномена, так в клетках её тела находится только 4 пары хромосом и имеет место высокая скорость размножения (в течение года можно исследовать более 20-ти поколений). Итак, сцепленными признаками называются признаки, которые контролируются генами, расположенными в одной хромосоме. Естественно, что они передаются вместе в случаях полного сцепления (закон Моргана). Полное сцепление встречается редко, обычно – неполное, из-за влияния кроссинговера (перекрещивания и обмена участками гомологичных хромосом в процессе мейоза). То есть, гены одной хромосомы переходят в другую, гомологичную ей.
Частота кроссинговера зависит от расстояния между генами. Чем ближе друг к другу расположены гены в хромосоме, тем сильнее между ними сцепление и тем реже происходит их расхождение при кроссинговере, и, наоборот, чем дальше друг от друга отстоят гены, тем слабее сцепление между ними и тем чаще возможно его нарушение.

На рисунке 1 слева : расстояние между генами А и В маленькое, вероятность разрыва хроматиды именно между А и В невелика, поэтому сцепление полное, хромосомы в гаметах идентичны родительским (два типа), других вариантов не появляется.
На рисунке 1 справа: расстояние между генами А и В большое, повышается вероятность разрыва хроматиды и последующего воссоединения крест-накрест именно между А и В , поэтому сцепление неполное, хромосомы в гаметах образуются четырёх типов – 2 идентичные родительским (некроссоверные) + 2 кроссоверных варианта.
Количество разных типов гамет будет зависеть от частоты кроссинговера или расстояния между анализируемыми генами. Расстояние между генами исчисляется в морганидах – единицах расстояния между генами, находящимися в одной хромосоме. 1 морганида соответствует 1 % кроссинговера. Такая зависимость между расстояниями и частотой кроссинговера прослеживается только до 50 морганид. Частота кроссинговера между определенной парой генов – довольно постоянная величина (хотя радиация, химические вещества, гормоны, лекарства влияют на нее; например, высокая температура стимулирует кроссинговер).
Пример, основанный на опытах Моргана

Гены, расположенные в одной хромосоме, наследуются совместно. Фенотипы А – серое тело, нормальные крылья (повторяет материнскую форму). Б – тёмное тело, короткие крылья (повторяет отцовскую форму). В – серое тело, короткие крылья (отличается от родителей). Г – тёмное тело, нормальные крылья (отличается от родителей). В и Г получены в результате кроссинговера в мейозе.
Если скрестить мушку дрозофилу, имеющую серое тело и нормальные крылья (на рисунке самка), с мушкой, обладающей тёмной окраской и зачаточными (короткими) крыльями (на рисунке самец), то в первом поколении гибридов все мухи будут серыми с нормальными крыльями (А). Это гетерозиготы по двум парам аллельных генов, причём ген, определяющий серую окраску брюшка, доминирует над тёмной окраской, а ген, обусловливающий развитие нормальных крыльев, - доминирует над геном недоразвитых крыльев.
При анализирующем скрещивании гибрида F 1 с гомозиготной рецессивной дрозофилой (Б) подавляющее большинство потомков F 2 будет сходно с родительскими формами.
Это происходит потому, что гены, отвечающие за серое тело и нормальные крылья– сцепленные гены, также как и гены, отвечающие за тёмное тело и короткие крылья, т. е. они находятся в одной хромосоме. наследование сцепленных генов называют сцепленным наследованием.
Сцепление может нарушаться. Это доказывает наличие особей В и Г на рисунке, т. е. если бы сцепление не нарушалось, то этих особей бы не существовало, однако они есть. Это происходит в результате кроссинговера, который и нарушает сцепленность этих генов.

На рисунке 3 опыт Моргана отображен подробно. Несцепленное наследование: два гена находятся в разных хромосомах, гетерозигота с равной вероятностью дает четыре типа гамет:

Сцепленное наследование : два гена находятся в одной хромосоме.
а) При полном сцеплении гетерозигота дает только два типа гамет.
б) При неполном сцеплении гетрозигота дает четыре типа гамет, но не с равной вероятностью.

На вышесказанном строится хромосомная теория наследственности Моргана:
1. Гены находятся в хромосомах и расположены в линейной последовательности на определенных расстояниях друг от друга.
2. Гены, расположенные в одной хромосоме, составляют группу сцепления. Число групп сцепления равно гаплоидному числу хромосом. Признаки, гены которых находятся в одной хромосоме, наследуются сцепленно (т. е. в тех же сочетаниях, в которых они были в хромосомах исходных родительских форм).
3. Новые сочетания генов, расположенных в одной паре хромосом, могут возникать в результате кроссинговера в процессе мейоза. Частота кроссинговера зависит от расстояния между генами.
4. Учитывая линейное расположение генов в хромосоме и частоту кроссинговера как показателя расстояния между генами, можно построить карты хромосом. За единицу расстояния между генами принята частота кроссинговера равная 1 % (морганида, сантиморган, сМ).
Задачи на нахождение расстояния между генами
Решение задачи на определение вероятности рождения здоровый и больных детей
Наследование признаков, сцепленных с полом
Сцепленными с полом называются признаки, гены которых расположены не в аутосоме (неполовой хромосоме), а в гетеросоме (половой хромосоме). Схема решения задач на наследование признаков, сцепленных с полом, иная, чем на аутосомное моногибридное скрещивание. В случае, если ген сцеплен с Х-хромосомой, он может передаваться от отца только дочерям, а от матери в равной степени и дочерям, и сыновьям. Если ген сцеплен с Х-хромосомой и является рецессивным, то у самки он проявляется только в гомозиготном состоянии. У самцов второй Х-хромосомы нет, поэтому такой ген проявляется всегда.
При решении задач этого типа используются не символы генов (А, а, В, b), как при аутосомном наследовании, а символы половых хромосом X, Y с указанием локализованных в них генов (X А , X а ).
Аномалии, сцепленные с полом, чаще контролируются рецессивными генами, локализованы в Х-хромосоме и проявляются при генотипе ХY (т. е. у самцов млекопитающих и самок птиц).
Выше были рассмотрены примеры, где ген, сцепленный с полом, располагался в Х-хромосоме, но есть гены, локализованные в Y-хромосоме. У видов, у которых мужской пол гетерогаметен, этот ген может передаваться только самцам. У человека ген одного из видов синдактилии, выражающейся в образовании перепонки между 2 и 3 пальцами на ноге, локализован на Y-хромосоме, поэтому синдактилия возникает только у мужчин. Известна еще одна аномалия – гипертрихоз края ушной раковины (ряды волос на ухе), передающиеся по такому же механизму. В изучаемой семье с этой аномалией она передавалась в пяти поколениях по мужской линии.
Хромосомная теория наследственности. Сцепленное наследование . Убедительные доказательства того, что гены располагаются в хромосомах, были получены в 1910 г. американским генетиком, будущим лауреатом Нобелевской премии (1933 г.) Т. Морганом. Многочисленные эксперименты Моргана и его сотрудников привели к ряду важнейших открытий, которые легли в основу хромосомной теории наследственности. Одно из положений этой теории можно сформулировать следующим образом: гены расположены в хромосомах в линейном порядке и занимают определенные участки — локусы, причем аллельные гены находятся в одинаковых локусах гомологичных хромосом.
Закон независимого наследования (третий закон Менделя) справедлив только в том случае, если неаллельные гены находятся в разных парах хромосом. Однако количество генов у живых организмов значительно больше числа хромосом. Например, у человека около 25 тыс. генов, а количество хромосом — 23 пары (2n = 46), у плодовой мушки дрозофилы приблизительно 14 тыс. генов и всего 4 пары хромосом (2n = 8). Следовательно, каждая хромосома содержит множество генов. Будут ли гены, локализованные в одной хромосоме, наследоваться независимо? Очевидно, что нет.
Гены, расположенные в одной хромосоме, образуют группу сцепления и наследуются вместе. Такое совместное наследование генов Т. Морган назвал сцепленным наследованием (в отличие от независимого). Гомологичные хромосомы каждой пары содержат гены, контролирующие одни и те же признаки, поэтому количество групп сцепления равно числу пар хромосом. Например, у человека 23 группы сцепления, а у дрозофилы — 4.
Вам известно, что при независимом наследовании дигетерозиготная особь, например , образует четыре типа гамет в равном соотношении, т. е. по 25 %: , , и . Это обусловлено тем, что неаллельные гены находятся в разных парах хромосом. Если же они расположены в гомологичных хромосомах, следовало бы ожидать, что дигетерозигота будет производить лишь два типа гамет: 50 % и 50 % . Обратите внимание на то, что сцепленные гены записываются в одну хромосому.
Однако Т. Морган обнаружил, что в большинстве случаев дигетерозиготные особи образуют не два типа гамет, а четыре. Кроме ожидаемых и , формируются также гаметы с новыми комбинациями генов: и , но в меньшем процентном соотношении, чем при независимом наследовании. Рассмотрим один из опытов, в котором Т. Морган изучал наследование сцепленных генов у дрозофилы.

*Дрозофила обыкновенная, или плодовая, — вид насекомых отряда Двукрылые (рис. 34.1). Особи, как правило, имеют желто-коричневое брюшко с темными поперечными кольцами и глаза красного цвета. В природе эти мушки питаются опавшими фруктами, соком растений, гниющими растительными остатками. В XX в. дрозофила стала одним из самых распространенных объектов исследований в области генетики и биологии индивидуального развития. Удобство ее использования обусловлено рядом причин. Так, дрозофилы имеют множество отчетливо различимых альтернативных признаков, небольшие размеры (обычно около 2—2,5 мм), легко разводятся в лабораторных условиях и обладают высокой плодовитостью — от каждого скрещивания можно получить до нескольких сотен потомков. Важно также и то, что время развития особи от откладки яйца до выхода половозрелой мушки из куколки при 25 °C занимает всего 10 дней.*
Чистую линию дрозофил, имеющих серое тело и нормальные (длинные) крылья, скрестили с чистой линией, особи которой имели черное тело и зачаточные крылья (рис. 34.2). Полученные гибриды первого поколения в соответствии с первым законом Менделя были единообразными — серыми с нормально развитыми крыльями. Следовательно, у дрозофилы серое тело (А) полностью доминирует над черным (а), а нормальные крылья (В) — над зачаточными (b). Все гибриды первого поколения — дигетерозиготы.
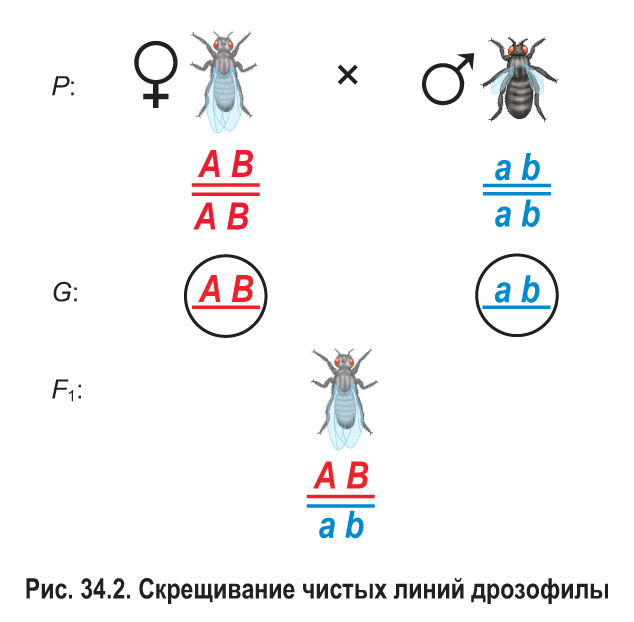
Затем было проведено анализирующее скрещивание (рис. 34.3). Дигетерозиготную самку из гибридного поколения скрестили с рецессивным дигомозиготным самцом (черное тело и зачаточные крылья). В потомстве было получено по 41,5 % особей с серым телом, нормальными крыльями и черным телом, зачаточными крыльями, а также по 8,5 % мух с серым телом, зачаточными крыльями и черным телом, нормальными крыльями.
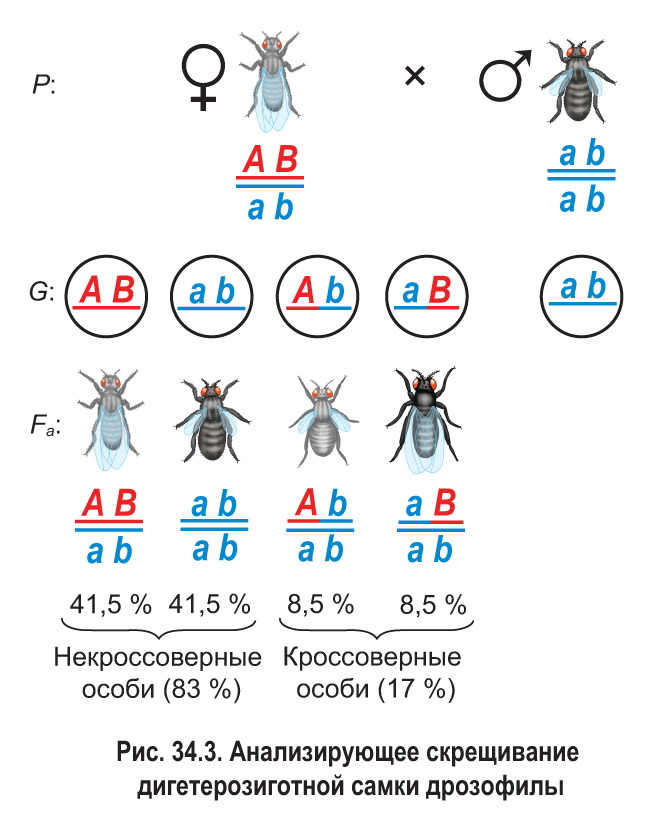
Если бы гены, определяющие цвет тела и развитие крыльев, находились в разных парах хромосом, соотношение фенотипических классов было бы равным — по 25 %. Но этого не наблюдалось, значит, гены находятся в гомологичных хромосомах и наследуются сцепленно.
Несмотря на сцепление генов, самка производила не два, а четыре типа гамет. Однако гамет с исходными сочетаниями сцепленных генов формировалось намного больше ( и вместе составили 83 %), чем с новыми их комбинациями (сумма и равна 17 %).
Было выяснено, что причиной появления хромосом с новыми комбинациями родительских генов является кроссинговер. Вы знаете, что этот процесс происходит в профазе I мейоза и представляет собой обмен соответствующими участками между гомологичными хромосомами. Таким образом, кроссинговер препятствует полному (абсолютному) сцеплению генов. Гаметы , которые образуются в результате кроссинговера, и особи, которые развиваются при участии таких гамет, называются кроссоверными или рекомбинантными. В рассмотренном эксперименте гаметы и являлись кроссоверными, а гаметы и — некроссоверными (см. рис. 34.3).
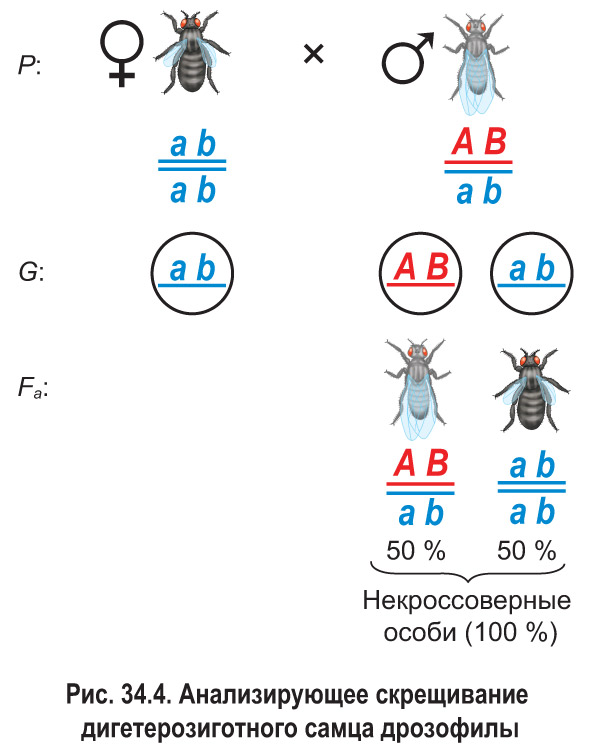
*Т. Морган подвергал анализирующему скрещиванию не только дигетерозиготных самок дрозофилы, но и самцов, т. е. проводил реципрóкные скрещивания. Так называют пару скрещиваний, в одном из которых организмы с исследуемыми признаками (или признаком) используются в качестве материнских, а в другом — в качестве отцовских. Оказалось, что реципрокные анализирующие скрещивания дигетерозиготных особей дрозофилы дают разные результаты.
В потомстве дигетерозиготного самца наблюдалось лишь два фенотипических класса (рис. 34.4). Половину потомков составляли особи с серым телом и нормальными крыльями, вторую половину — особи с черным телом и зачаточными крыльями. Это свидетельствовало об отсутствии кроссинговера между сцепленными генами или, иначе говоря, о полном (абсолютном) сцеплении генов. Выяснилось, что у самцов дрозофилы в ходе мейоза действительно не происходит кроссинговер между гомологичными хромосомами. Впоследствии подобное явление было обнаружено у самцов некоторых других видов двукрылых, а также у самок тутового шелкопряда. Однако полное сцепление генов наблюдается в природе очень редко, как исключение из общего правила.*
Кроссинговер между сцепленными генами происходит с определенной вероятностью (частотой). Для расчета частоты кроссинговера (rf, от англ. recombination frequency — частота рекомбинации) можно пользоваться следующей формулой:
Таким образом, между генами А и В, контролирующими цвет тела и длину крыльев дрозофилы, кроссинговер происходит с частотой: rfAB = 17 %.
Дальнейшие исследования, проведенные Т. Морганом и сотрудниками его лаборатории, показали, что частота кроссинговера пропорциональна расстоянию между генами, расположенными в одной хромосоме. Чем больше расстояние между сцепленными генами, тем чаще между ними происходит кроссинговер. И наоборот, чем ближе друг к другу расположены гены, тем меньше частота кроссинговера между ними. Чем объясняется эта закономерность?
В профазе I мейоза при конъюгации гомологичных хромосом взаимный обмен участками между хроматидами осуществляется произвольно, в любой их точке. Рассмотрим рисунок 34.5.
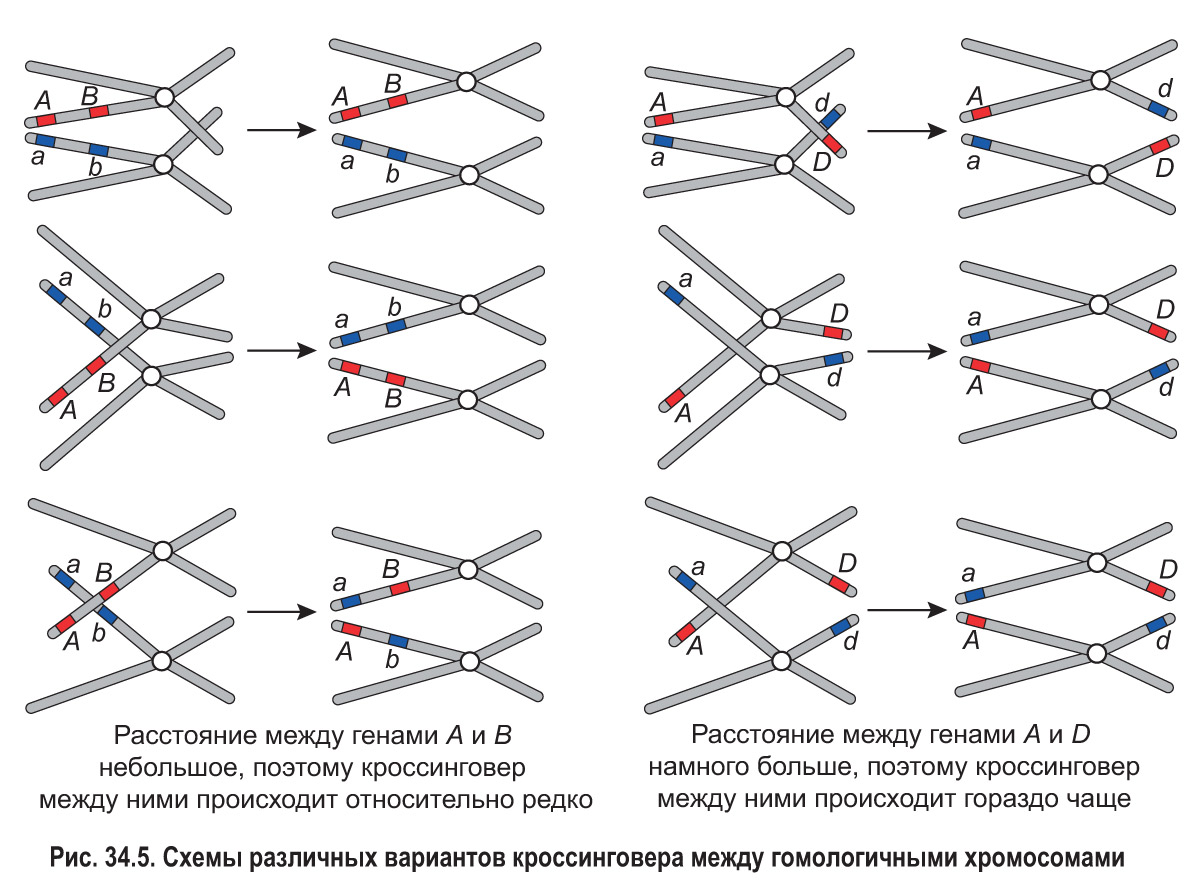
Гены А и В (или а и b) находятся сравнительно близко друг к другу. Вероятность того, что обмен произойдет именно на участке, разделяющем эти гены, невелика. Гены А и D (или а и d) располагаются на значительном расстоянии друг от друга. Поэтому вероятность того, что хроматиды совершат обмен в какой-либо точке между этими генами, намного выше. Значит, чем больше расстояние между генами, тем чаще они разделяются при кроссинговере.
Таким образом, частота кроссинговера позволяет судить о расстоянии между генами. В честь Т. Моргана единица измерения расстояния между генами получила название морганида или, что то же самое, сантиморган (сМ).
Морганида (сантиморган, сМ) — это генетическое расстояние, на котором кроссинговер происходит с вероятностью 1 %.
Кроссинговер играет очень важную биологическую роль. В результате этого процесса у потомков возникают новые комбинации родительских генов, что повышает генетическое разнообразие организмов и расширяет возможности их адаптации к различным условиям окружающей среды.
В 1906 году У. Бэтсон и Р. Пеннет, проводя скрещивание растений душистого горошка и анализируя наследование формы пыльцы и окраски цветков, обнаружили, что эти признаки не дают независимого распределения в потомстве, гибриды всегда повторяли признаки родительских форм. Стало ясно, что не для всех признаков характерно независимое распределение в потомстве и свободное комбинирование.
Каждый организм имеет огромное количество признаков, а число хромосом невелико. Следовательно, каждая хромосома несет не один ген, а целую группу генов, отвечающих за развитие разных признаков. Изучением наследования признаков, гены которых локализованы в одной хромосоме, занимался Т. Морган. Если Мендель проводил свои опыты на горохе, то для Моргана основным объектом стала плодовая мушка дрозофила.
Дрозофила каждые две недели при температуре 25 °С дает многочисленное потомство. Самец и самка внешне хорошо различимы — у самца брюшко меньше и темнее. Они имеют всего 8 хромосом в диплоидном наборе, достаточно легко размножаются в пробирках на недорогой питательной среде.
Скрещивая мушку дрозофилу с серым телом и нормальными крыльями с мушкой, имеющей темную окраску тела и зачаточные крылья, в первом поколении Морган получал гибриды, имеющие серое тело и нормальные крылья (ген, определяющий серую окраску брюшка, доминирует над темной окраской, а ген, обусловливающий развитие нормальных крыльев, — над геном недоразвитых). При проведении анализирующего скрещивания самки F1 с самцом, имевшим рецессивные признаки, теоретически ожидалось получить потомство с комбинациями этих признаков в соотношении 1:1:1:1. Однако в потомстве явно преобладали особи с признаками родительских форм (41,5% — серые длиннокрылые и 41,5% — черные с зачаточными крыльями), и лишь незначительная часть мушек имела иное, чем у родителей, сочетание признаков (8,5% — черные длиннокрылые и 8,5% — серые с зачаточными крыльями). Такие результаты могли быть получены только в том случае, если гены, отвечающие за окраску тела и форму крыльев, находятся в одной хромосоме.

1 — некроссоверные гаметы; 2 — кроссоверные гаметы.
Если гены окраски тела и формы крыльев локализованы в одной хромосоме, то при данном скрещивании должны были получиться две группы особей, повторяющие признаки родительских форм, так как материнский организм должен образовывать гаметы только двух типов — АВ и аb , а отцовский — один тип — аb . Следовательно, в потомстве должны образовываться две группы особей, имеющих генотип ААВВ и ааbb . Однако в потомстве появляются особи (пусть и в незначительном количестве) с перекомбинированными признаками, то есть имеющие генотип Ааbb и ааВb . Для того, чтобы объяснить это, необходимо вспомнить механизм образования половых клеток — мейоз. В профазе первого мейотического деления гомологичные хромосомы конъюгируют, и в этот момент между ними может произойти обмен участками. В результате кроссинговера в некоторых клетках происходит обмен участками хромосом между генами А и В , появляются гаметы Аb и аВ , и, как следствие, в потомстве образуются четыре группы фенотипов, как при свободном комбинировании генов. Но, поскольку кроссинговер происходит при образовании небольшой части гамет, числовое соотношение фенотипов не соответствует соотношению 1:1:1:1.
Группа сцепления — гены, локализованные в одной хромосоме и наследующиеся совместно. Количество групп сцепления соответствует гаплоидному набору хромосом.
Сцепленное наследование — наследование признаков, гены которых локализованы в одной хромосоме. Сила сцепления между генами зависит от расстояния между ними: чем дальше гены располагаются друг от друга, тем выше частота кроссинговера и наоборот. Полное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются так близко друг к другу, что кроссинговер между ними становится невозможным. Неполное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются на некотором расстоянии друг от друга, что делает возможным кроссинговер между ними.
Независимое наследование — наследование признаков, гены которых локализованы в разных парах гомологичных хромосом.
Некроссоверные гаметы — гаметы, в процессе образования которых кроссинговер не произошел.
| Образуются гаметы: |
Кроссоверные гаметы — гаметы, в процессе образования которых произошел кроссинговер. Как правило кроссоверные гаметы составляют небольшую часть от всего количества гамет.
| Образуются гаметы: |
Нерекомбинанты — гибридные особи, у которых такое же сочетание признаков, как и у родителей.
Рекомбинанты — гибридные особи, имеющие иное сочетание признаков, чем у родителей.
Расстояние между генами измеряется в морганидах — условных единицах, соответствующих проценту кроссоверных гамет или проценту рекомбинантов. Например, расстояние между генами серой окраски тела и длинных крыльев (также черной окраски тела и зачаточных крыльев) у дрозофилы равно 17%, или 17 морганидам.
У дигетерозигот доминантные гены могут располагаться или в одной хромосоме (цис-фаза), или в разных (транс-фаза).

1 — Механизм цис-фазы (некроссоверные гаметы); 2 — механизм транс-фазы (некроссоверные гаметы).
Результатом исследований Т. Моргана стало создание им хромосомной теории наследственности:
- гены располагаются в хромосомах; различные хромосомы содержат неодинаковое число генов; набор генов каждой из негомологичных хромосом уникален;
- каждый ген имеет определенное место (локус) в хромосоме; в идентичных локусах гомологичных хромосом находятся аллельные гены;
- гены расположены в хромосомах в определенной линейной последовательности;
- гены, локализованные в одной хромосоме, наследуются совместно, образуя группу сцепления; число групп сцепления равно гаплоидному набору хромосом и постоянно для каждого вида организмов;
- сцепление генов может нарушаться в процессе кроссинговера, что приводит к образованию рекомбинантных хромосом; частота кроссинговера зависит от расстояния между генами: чем больше расстояние, тем больше величина кроссинговера;
- каждый вид имеет характерный только для него набор хромосом — кариотип.
Сцепленными являются признаки, гены которых локализованы в одной хромосоме. В случае полного сцепления они передаются совместно и у дигетерозигот образуется лишь 2 вида гамет. Если гены сцеплены относительно, то в мейозе происходит кроссинговер и образуются кроссоверные и некроссоверные гаметы. У дигетерозигот формируется 2 вида некроссоверных и 2 вида кроссоверных гамет. Число разных типов гамет зависит от расстояния между анализируемыми генами, которое выражается в морганидах или процентах кроссинговера.
Образование гамет при сцепленном наследовании
Задача 87. У дрозофилы гены А, а и В, в расположены в одной паре аутосом, причём гены А и в, а также а и В абсолютно сцеплены. Какие виды гамет и в каком соотношении образует дрозофила с генотипом АаВв?
Решение: Укажем схему расположения генов в паре хромосом:А в
Так как сцеплены гены А и в, они расположены в одной хромосоме, а гены а и В во второй гомологичной хромосоме. Поскольку гены сцеплены абсолютно, кроссинговер не происходит, значит образуется 2 вида гамет Ав и аВ, они образуются в равных соотношениях, т.е. по 50%.
Ответ: дрозофила с генотипом АаВв образует два вида гамет Ав и аВ по 50%.
Задача 88. Гены М и N, а также их рецессивные аллели m и n расположены в одной паре гомологичных хромосом. Расстояние между генами М и N, а также m и n сцеплены не абсолютно и расстояние между ними равно 20 морганидам, образуется 20% кроссоверных гамет и 100 – 20 = 80% некроссоверных гамет. Некроссоверных гамет образуется 2 вида: MN и mn – всего 80%, на каждый вид по 40%. Кроссоверные гаметы – тоже 2 вида: Mn и mN – всего 20%, на каждый вид 10%.
Ответ: организм с генотипом MmNn образует 4 вида гамет, из них 2 вида некроссоверные: MN и mn – по 40% и 2 вида кроссоверные: Mn и mN – по 10%.
Задача 89. У дрозофилы гены С и Д и их рецессивные аллели локализованы в одной паре гомологичных хромосом, причём расстояние между ними равно 40 морганидам. Гены Е, е расположены в другой паре хромосом. Какие типы гамет и в каком соотношении образует дрозофила с генотипом СсДdЕе?
Решение: Приведём схему расположения генов в хромосомах:
С Д Е Расстояние между генами С и Д, а также с и d указывает
с d е на количество кроссоверных гамет (40%), количество некроссоверных гамет равно 100 – 40 = 60%. Так как в данной задаче анализируются гены, расположенные в двух парах гомологичных хромосомах, а хромосомы в мейозе свободно комбинируются, образуется 4 вида некроссоверных гамет и 4 вида кроссоверных гамет. Некроссоверные гаметы: СДЕ, СДе, сdЕ, сdе – всего 60%, на каждый вид по 15%. Кроссоверные гаметы: СdЕ, Сdе, сДЕ, сДе – всего 40%, на каждый вид по 10%.
Ответ: дрозофила с генотипом СсД Ее образует 8 видов гамет, из них 4 вида некроссоверные: СДЕ, СДе, сdЕ, сdе по 15% и 4 вида кроссоверные: СdЕ, Сdе, сДЕ, сДе по 10%.
Задача 90.Гены Е и F и их рецессивные аллели с и f расположены в одной паре хромосом и абсолютно сцеплены. Какие виды гамет и в каком соотношении образует организм с генотипом ЕеFf?
Задача 91. Гены С, с и Д, d локализованы в одной паре гомологичных хромосом, причем расстояние между генами С и d, а также с и Д равно 10 морганидам. Сколько и какие виды гамет образует организм с генотипом СсДd?
Задача 92. У дрозофилы гены А и В и их рецессивные аллели а и в находятся в одной паре хромосом и сцеплены абсолютно. Гены С и с локализованы в другой паре хромосом. Сколько и какие виды гамет образуются у дрозофилы с генотипом АаВвСс?
Задача 93. Гены А и а расположены в одной паре гомологичных хромосом, а а гены Д и Е и их рецессивные аллели – в другой. Расстояние между генами Д и е, а также d и Е равно 20 морганидам. Какие виды гамет и в каком соотношении образует организм с генотипом АаДdЕе?
Задача 94. Гены А, а и В, в находятся в одной паре хромосом, причем гены А и В, а также а и в абсолютно сцеплены. Гены С и Д и их рецессивные аллели с и d расположены в другой паре хромосом. Гены С и d, а также с и Д тоже полностью сцеплены. Сколько и какие виды гамет образует организм с генотипом АаВвСсДd?
Задача 95. У дрозофилы гены А и В и их рецессивные аллели а и в расположены в одной паре гомологичных хромосом. Расстояние между генами А и В, а также а и в равно 24 морганидам. Гены Е, е и F, f находятся в другой паре хромосом, причём гены Е и F, а также е и f полностью сцеплены. Какие виды гамет и в каком соотношении образует организм с генотипом АаВвЕеFf?
Задача 96.Гены В, в и С, с находятся в одной паре хромосом, причём гены В и с, а также в и С абсолютно сцеплены. Гены Д и Е и их рецессивные аллели расположены в другой паре хромосом, причём расстояние между генами Д и е, а также d и Е равно 12 морганидам. Сколько и какие виды гамет образует организм с генотипом ВвСсДdЕе?
Читайте также:
- Мошенничество какой состав формальный или материальный состав
- В каких случаях должностные лица органов гпн применяют уголовную ответственность
- Радио какие есть должности
- Какой институт политической системы обладает легальным правом на применение силы политические партии
- Муниципальное право как отрасль российского права сформировалось в веке