
Какой тип взаимодействия двух генов лежит в основе наследования окраски клубней и цветков картофеля
Обновлено: 17.05.2024
Теперь обратимся к проблеме взаимодействия неаллельных генов. Если развитие признака контролируется более чем одной парой генов, то это означает, что он находится под полигенным контролем. Установлено несколько основных типов взаимодействия генов: комплементарность, эпистаз, полимерия и плейотропия.
Первый случай неаллельного взаимодействия был описан в качестве примера отклонения от законов Менделя английскими учеными У. Бетсоном и Р. Пеннетом в 1904 г. при изучении наследования формы гребня у кур. Различные породы кур характеризуются разной формой гребня. Виандотты имеют низкий, правильный, покрытый сосочками гребень, известный под названием “розовидного”. Брамы и некоторые бойцовые куры обладают узким и высоким гребнем с тремя продольными возвышениями — “гороховидным”. Леггорны имеют простой или листовидный гребень, состоящий из одной вертикальной пластинки. Гибридологический анализ показал, что простой гребень ведет себя как полностью рецессивный признак по отношению к розовидному и гороховидному. Расщепление в F2 соответствует формуле 3 : 1. При скрещивании же между собой рас с розовидным и гороховидным гребнем у гибридов первого поколения развивается совершенно новая форма гребня, напоминающая половинку ядра грецкого ореха, в связи с чем гребень был назван “ореховидным”. При анализе второго поколения было установлено, что соотношение разных форм гребня в F2 соответствует формуле 9 : 3 : 3 : 1, что указывало на дигибридный характер скрещивания. Была разработана схема скрещивания, объясняющая механизм наследования этого признака.
В определении формы гребня у кур принимают участие два неаллельных гена. Доминантный ген R контролирует развитие розовидного гребня, а доминантный ген P — гороховидного. Комбинация рецессивных аллелей этих генов rrpp вызывает развитие простого гребня. Ореховидный гребень развивается при наличии в генотипе обоих доминантных генов.
Наследование формы гребня у кур можно отнести к комплементарному взаимодействию неаллельных генов. Комплементарными, или дополнительными, считаются гены, которые при совместном действии в генотипе в гомо- или гетерозиготном состоянии обусловливают развитие нового признака. Действие же каждого из генов в отдельности воспроизводит признак одного из родителей.

Схема, иллюстрирующая взаимодействие неаллельных генов,
определяющих форму гребня у кур
Наследование генов, определяющих форму гребня у кур, полностью укладывается в схему дигибридного скрещивания, так как они ведут себя при распределении независимо. Отличие от обычного дигибридного скрещивания проявляется только на уровне фенотипа и сводится к следующему:
- Гибриды F1 не похожи ни на одного из родителей и обладают новым признаком;
- В F2 появляются два новых фенотипических класса, которые являются результатом взаимодействия либо доминантных (ореховидный гребень), либо рецессивных (простой гребень) аллелей двух независимых генов.
Механизм комплементарного взаимодействия подробно изучен на примере наследования окраски глаз у дрозофилы. Красная окраска глаз у мух дикого типа определяется одновременным синтезом двух пигментов — бурого и ярко-красного, каждый из которых контролируется доминантным геном. Мутации, затрагивающие структуру этих генов, блокируют синтез либо того, либо другого пигмента. Так, рецессивная мутация brown (ген находится во 2-й хромосоме) блокирует синтез ярко-красного пигмента, в связи с чем у гомозигот по этой мутации бурые глаза. Рецессивная мутация scarlet (ген располагается в 3-й хромосоме) нарушает синтез бурого пигмента, и поэтому гомозиготы stst имеют ярко-красные глаза. При одновременном присутствии в генотипе обоих мутантных генов в гомозиготном состоянии не вырабатываются оба пигмента и глаза у мух белые.
В описанных примерах комплементарного взаимодействия неаллельных генов формула расщепления по фенотипу в F2 соответствует 9 : 3 : 3 : 1. Такое расщепление наблюдается в том случае, если взаимодействующие гены по отдельности имеют неодинаковое фенотипическое проявление и оно не совпадает с фенотипом гомозиготного рецессива. Если это условие не соблюдается, в F2 имеют место иные соотношения фенотипов.
Например, при скрещивании двух разновидностей фигурной тыквы со сферической формой плода гибриды первого поколения обладают новым признаком — плоскими или дисковидными плодами. При скрещивании гибридов между собой в F2 наблюдается расщепление в соотношении 9 дисковидных : 6 сферических : 1 удлиненная.

Схема наследования формы плода у тыкв
Анализ схемы показывает, что в определении формы плода принимают участие два неаллельных гена с одинаковым фенотипическим проявлением (сферическая форма). Взаимодействие доминантных аллелей этих генов дает дисковидную форму, взаимодействие рецессивных аллелей — удлиненную.
Еще один пример комплементарного взаимодействия дает наследование окраски шерсти у мышей. Дикая серая окраска определяется взаимодействием двух доминантных генов. Ген А отвечает за присутствие пигмента, а ген В — за его неравномерное распределение. Если в генотипе присутствует только ген А (А-bb), то мыши равномерно окрашены в черный цвет. Если присутствует только ген В (ааВ-), то пигмент не вырабатывается и мыши оказываются неокрашенными, так же как и гомозиготный рецессив ааbb. Такое действие генов приводит к тому, что в F2 расщепление по фенотипу соответствует формуле 9 : 3 : 4.
Комплементарное взаимодействие описано также при наследовании окраски цветов у душистого горошка. Большая часть сортов этого растения имеет пурпурные цветы с фиолетовыми крыльями, которые характерны для дикой сицилийской расы, но есть также сорта с белой окраской. Скрещивая растения с пурпурной окраской цветов с растениями с белыми цветами Бетсон и Пеннет установили, что пурпурная окраска цветов полностью доминирует над белой, и в F2 наблюдается соотношение 3 : 1. Но в одном случае от скрещивания двух белых растений получилось потомство, состоящее только из растений с окрашенными цветами. При самоопылении растений F1 было получено потомство, состоящее из двух фенотипических классов: с окрашенными и неокрашенными цветами в соотношении 9/16 : 7/16.
Полученные результаты объясняются комплементарным взаимодействием двух пар неаллельных генов, доминантные аллели которых (С и Р) в отдельности не способны обеспечить развитие пурпурной окраски, так же как и их рецессивные аллели (ссрр). Окраска проявляется только при наличии в генотипе обоих доминантных генов, взаимодействие которых обеспечивает синтез пигмента.
В приведенном примере формула расщепления в F2 — 9 : 7 обусловлена отсутствием у доминантных аллелей обоих генов собственного фенотипического проявления. Однако такой же результат получается и в том случае, если взаимодействующие доминантные гены имеют одинаковое фенотипическое проявление. Например, при скрещивании двух сортов кукурузы с фиолетовой окраской зерновок в F1 все гибриды имеют желтые зерновки, а в F2 наблюдается расщепление 9/16 желт. : 7/16 фиол.
Эпистаз — другой тип неаллельного взаимодействия, при котором происходит подавление действия одного гена другим неаллельным ему геном. Ген, который препятствует проявлению другого гена, называется эпистатичным, или супрессором, а тот, чье действие подавляется, гипостатичным. В качестве эпистатичного гена может выступать как доминантный, так и рецессивный ген (соответственно доминантный и рецессивный эпистаз).
Примером доминантного эпистаза служит наследование окраски шерсти у лошадей и окраски плодов у тыквы. Схема наследования этих двух признаков абсолютно одинаковая.
Схема наследования окраски шерсти у лошадей
F2
| CB | Cb | cB | cb | |
| CB | CCBB сер. | CCBB сер. | CcBB сер. | CcBb сер. |
| Cb | CCBb сер. | CCbb сер. | CcBb сер. | Ccbb сер. |
| cB | CcBB сер. | CcBb сер. | ccBB черн. | ccBb черн. |
| cb | CcBb сер. | Ccbb сер. | ccBb черн. | ccbb рыж. |
Из схемы видно, что доминантный ген серой окраски С является эпистатичным по отношению к доминантному гену В, который обусловливает черную окраску. В присутствии гена С ген В своего действия не проявляет, и поэтому гибриды F1 несут признак, определяемый эпистатичным геном. В F2 класс с обоими доминантными генами сливается по фенотипу (серая окраска) с классом, у которого представлен только эпистатичный ген (12/16). Черная окраска проявляется у 3/16 гибридных потомков, в генотипе которых отсутствует эпистатичный ген. В случае гомозиготного рецессива отсутствие гена-супрессора позволяет проявиться рецессивному гену с, который вызывает развитие рыжей окраски.

Схема наследования окраски оперения у кур
Доминантный эпистаз описан также при наследовании окраски пера у кур. Белый цвет оперенья у кур породы леггорнов доминирует над окрашенным черных, рябых и других цветных пород. Однако белая окраска других пород (например, плимутроков) рецессивна по отношению к цветному оперению. Скрещивания между особями с доминантной белой окраской и особями с рецессивной белой окраской в F1 дают белое потомство. В F2 наблюдается расщепление в соотношении 13 : 3.
Анализ схемы показывает, что в определении окраски пера у кур принимают участие две пары неаллельных генов. Доминантный ген одной пары (I) является эпистатичным по отношению к доминантному гену другой пары, вызывающему развитие окраски (C). В связи с этим окрашенное оперение имеют только те особи, в генотипе которых присутствует ген С, но отсутствует эпистатичный ген I. У рецессивных гомозигот ссii отсутствует эпистатичный ген, но у них нет гена, который обеспечивает выработку пигмента (C), поэтому они имеют белую окраску.
В качестве примера рецессивного эпистаза можно рассмотреть ситуацию с геном альбинизма у животных (см. выше схему наследования окраски шерсти у мышей). Присутствие в генотипе двух аллелей гена альбинизма (аа) не дает возможности проявиться доминантному гену окраски (B) — генотипы ааВ-.
Полимерный тип взаимодействия был впервые установлен Г. Нильсеном-Эле при изучении наследования окраски зерна у пшеницы. При скрещивании краснозерного сорта пшеницы с белозерным в первом поколении гибриды были окрашенными, но окраска была розовой. Во втором поколении только 1/16 часть потомства имела красную окраску зерна и 1/16 — белую, у остальных окраска была промежуточной с разной степенью выраженности признака (от бледно-розовой до темно-розовой). Анализ расщепления в F2 показал, что в определении окраски зерна участвуют две пары неаллельных генов, действие которых суммируется. Степень выраженности красной окраски зависит от количества доминантных генов в генотипе.
Полимерные гены принято обозначать одинаковыми буквами с добавлением индексов, в соответствии с числом неаллельных генов.
Действие доминантных генов в данном скрещивании является аддитивным, так как добавление любого из них усиливает развитие признака.
Описанный тип полимерии, при котором степень развития признака зависит от дозы доминантного гена, называется кумулятивным. Такой характер наследования обычен для количественных признаков, к которым следует отнести и окраску, т.к. ее интенсивность обусловлена количеством вырабатываемого пигмента. Если не учитывать степень выраженности окраски, то соотношение окрашенных и неокрашенных растений в F2 соответствует формуле 15 : 1.
Однако в некоторых случаях полимерия не сопровождается кумулятивным эффектом. В качестве примера можно привести наследование формы семян у пастушьей сумки. Скрещивание двух рас, одна из которых имеет треугольные плоды, а другая яйцевидные дает в первом поколении гибриды с треугольной формой плода, а во втором поколении наблюдается расщепление по этим двум признакам в соотношении 15 треуг. : 1 яйцев.
Данный случай наследования отличается от предыдущего только на фенотипическом уровне: отсутствие кумулятивного эффекта при увеличении дозы доминантных генов обусловливает одинаковую выраженность признака (треугольная форма плода) независимо от их количества в генотипе.
К взаимодействию неаллельных генов относят также явление плейотропии — множественного действия гена, влияния его на развитие нескольких признаков. Плейотропное действие генов является результатом серьезного нарушения обмена веществ, обусловленного мутантной структурой данного гена.
Так, например, ирландские коровы породы декстер отличаются от близкой по происхождению породы керри укороченными ногами и головой, но одновременно лучшими мясными качествами и способностью к откорму. При скрещивании коров и быков породы декстер 25% телят имеют признаки породы керри, 50% сходны с породой декстер, а в остальных 25% случаев наблюдаются выкидыши уродливых бульдогообразных телят. Генетический анализ позволил установить, что причиной гибели части потомства является переход в гомозиготное состояние доминантной мутации, вызывающей недоразвитие гипофиза. В гетерозиготе этот ген приводит к появлению доминантных признаков коротконогости, короткоголовости и повышенной способности к отложению жира. В гомозиготе этот ген имеет летальный эффект, т.е. в отношении гибели потомства он ведет себя как рецессивный ген.
Летальный эффект при переходе в гомозиготное состояние характерен для многих плейотропных мутаций. Так, у лисиц доминантные гены, контролирующие платиновую и беломордую окраски меха, не оказывающие летального действия в гетерозиготе, вызывают гибель гомозиготных зародышей на ранней стадии развития. Аналогичная ситуация имеет место при наследовании серой окраски шерсти у овец породы ширази и недоразвития чешуи у зеркального карпа. Летальный эффект мутаций приводит к тому, что животные этих пород могут быть только гетерозиготными и при внутрипородных скрещиваниях дают расщепление в соотношении 2 мутанта : 1 норма.
Схема наследования платиновой окраски у лис
F1
| A | a | |
| A | AA погибают | Aa платин. |
| a | Aa платин. | aa черн. |
Однако большинство летальных генов рецессивны, и гетерозиготные по ним особи имеют нормальный фенотип. О наличии у родителей таких генов можно судить по появлению в потомстве гомозиготных по ним уродов, абортусов и мертворожденных. Чаще всего подобное наблюдается в близкородственных скрещиваниях, где родители обладают сходными генотипами, и шансы перехода вредных мутаций в гомозиготное состояние достаточно высоки.
Плейотропные гены с летальным эффектом есть у дрозофилы. Так, доминантные гены Curly — загнутые вверх крылья, Star — звездчатые глаза, Notch — зазубренный край крыла и ряд других в гомозиготном состоянии вызывают гибель мух на ранних стадиях развития.
Известная рецессивная мутация white, впервые обнаруженная и изученная Т. Морганом, также имеет плейотропный эффект. В гомозиготном состоянии этот ген блокирует синтез глазных пигментов (белые глаза), снижает жизнеспособность и плодовитость мух и видоизменяет форму семенников у самцов.
У человека примером плейотропии служит болезнь Марфана (синдром паучьих пальцев, или арахнодактилия), которая вызывается доминантным геном, вызывающим усиленный рост пальцев. Одновременно он определяет аномалии хрусталика глаза и порок сердца. Болезнь протекает на фоне повышения интеллекта, в связи с чем ее называют болезнью великих людей. Ею страдали А. Линкольн, Н. Паганини.
Плейотропный эффект гена, по всей видимости, лежит в основе коррелятивной изменчивости, при которой изменение одного признака влечет за собой изменение других.
К взаимодействию неаллельных генов следует отнести также влияние генов-модификаторов, которые ослабляют или усиливают функцию основного структурного гена, контролирующего развитие признака. У дрозофилы известны гены-модификаторы, модифицирующие процесс жилкования крыльев. Известно не менее трех генов-модификаторов, влияющих на количество красного пигмента в волосе крупного рогатого скота, в результате чего окраска шерсти у разных пород колеблется от вишневой до палевой. У человека гены-модификаторы изменяют окраску глаз, усиливая или ослабляя ее интенсивность. Их действием объясняется разная окраска глаз у одного человека.
Существование явления взаимодействия генов привело к появлению таких понятий, как “генотипическая среда” и “генный баланс”. Под генотипической средой подразумевается то окружение, в которое попадает вновь возникающая мутация, т.е. весь комплекс генов, имеющихся в данном генотипе. Понятие “генный баланс” касается соотношения и взаимодействия между собой генов, влияющих на развитие признака. Обычно гены обозначают названием признака, возникающего при мутации. На самом же деле проявление этого признака часто является результатом нарушения функции гена под влиянием других генов (супрессоров, модификаторов и др.). Чем сложнее генетический контроль признака, чем больше генов участвуют в его развитии, тем выше наследственная изменчивость, так как мутация любого гена нарушает генный баланс и приводит к изменению признака. Следовательно, для нормального развития особи необходимо не только присутствие генов в генотипе, но и осуществление всего комплекса межаллельных и неаллельных взаимодействий.
Перейти к чтению других тем книги "Генетика и селекция. Теория. Задания. Ответы":
Задача 1
У душистого горошка окраска цветов проявляется только при наличии двух доминантных генов А и В. Если в генотипе имеется только один доминантный ген, то окраска не развивается. Какое потомство F1 и F2 получится от скрещивания растений с генотипами ААbb и ааВВ?
Решение:
1. Генотип исходных растений известен по условию задачи. Они гомозиготны и будут давать один тип гамет, которые можно объединить единственным образом. Потомство F1 будет единообразно по генотипу (АаВb) и фенотипу (розовые цветы):
Схема первого скрещивания:
2. Дигетерозиготные потомки F1 будут давать по 4 типа гамет, которые могут комбинироваться 16 способами. Построив решетку Пеннета, можно убедиться, что организмов, одновременно имеющих доминантные гены А и В (розовые цветы), будет примерно 9/16. То есть, в F2 будет наблюдаться расщепление по цвету в пропорции 9:7.
Схема второго скрещивания
Ответ:
В F1 все потомство будет единообразным и будет иметь розовую окраску цветов. В F2 9/16 растений будут иметь розовые, а 7/16 – белые цветы.
Задача 2
При скрещивании двух растений тыквы со сферической формой плодов получено потомство, имеющее только дисковидные плоды. При скрещивании этих гибридов между собой были получены растения с тремя типами плодов:
• 9 частей – с дисковидными плодами,
• 6 частей – со сферической формой плодов,
• 1 часть – с удлиненными плодами.
Какая закономерность наблюдается в данном случае? Каковы генотипы родителей и потомства?
Решение:
1. Потомство F1 отличалось от родителей и было единообразным, следовательно, оно является гетерозиготным, а исходные особи были гомозиготными.
Схема первого скрещивания:
2. Исходные особи должны отличаться по генотипу друг от друга, так как при одинаковом генотипе их потомство не отличалось бы от них. Это указывает на то, что в данном случае имеет место комплементарное взаимодействие генов. Генотип родительских особей – BBcc и bbCC, а потомства F1 – BbCc.
3. Таким образом, форма плодов определяется двумя парами генов, В и С. Если обе пары находятся в доминантном состоянии (генотипы BBCC, BbCc, BBCc или BbCC), то форма плодов дисковидная. Если в доминантном состоянии находится только одна пара генов (генотипы BBcc, Bbcc, bbCC или bbCc), – форма плодов сферическая.
4. Расщепление в потомстве F2 (9:6:1) отличается от расщепления 9:7, обычно наблюдаемого при комплементарном взаимодействии. 1/16 часть растений имела плоды удлиненной формы. Это может быть только в том случае, если генотип таких особей – bbсс, то есть оба гена находятся в рецессивном состоянии.
Схема второго скрещивания:
5. Анализ решетки Пеннета подтверждает полученные результаты.
Задача 3
У норки известно два рецессивных гена – р и i, гомозиготность по каждому из которых, или по обоим одновременно, обуславливает платиновую окраску меха. Дикая коричневая окраска получается при наличии обоих доминантных аллелей Р и I. При каком типе скрещивания двух платиновых норок все их потомство будет коричневым?
Решение:
Платиновые норки должны быть обязательно гомозиготны по каждому из двух рецессивных генов - р и i или по обоим одновременно, т.е. генотип особей должен быть —Ppii, ppIi, ppII, PPii или ppii. Дикую коричневую окраску будут иметь особи с генотипом PpIi. В случае, если при скрещивании двух платиновых норок всё их потомство будет коричневым, генотип одного родителя должен быть гомозиготен по рецессивному гену р, а по гену i – гетерозиготен; другой родитель, наоборот, гомозиготен по гену i и гетерозиготен по гену р. Тогда для скрещивания необходимо взять животных с генотипами ppII и PPii. Что подтверждается схемой скрещивания.
Ответ:
Для скрещивания необходимо взять животных с генотипами ppII и PPii.
Задача 4
У кукурузы нормальный рост определяется двумя доминантными неаллельными генами. Гомозиготность по рецессивным аллелям даже одной пары генов приводит к возникновению карликовых форм. При скрещивании двух карликовых растений кукурузы выросли гибриды нормальной высоты, а при скрещивании этих гибридов в их потомстве было получено 812 нормальных и 640 карликовых растений. Определить генотипы родителей и потомков.
Решение:
1. Так как при скрещивании двух карликовых растений было получено потомство нормальной высоты. Потомство F1 отличалось от родителей и было единообразным, следовательно, оно является гетерозиготным, а исходные особи были гомозиготными. Таким образом, родители имели генотип ААbb и aaBB, а потомство — AaBb.
Схема первого скрещивания:
2. Дигетерозиготные потомки F1 будут давать по 4 типа гамет, которые могут комбинироваться 16 способами. Построив решетку Пеннета, можно убедиться, что организмов, одновременно имеющих доминантные гены А и В (нормальный рост), будет примерно 9/16. То есть, в F2 будет наблюдаться расщепление по карликовому росту в пропорции 9:7.
Схема второго скрещивания:
3. в потомстве F2 наблюдается расщепление 9:7, характерное для комплементарного наследования.
Ответ:
Исходные организмы имели генотипы ААbb и ааВВ. Единообразное потомство F1 несло генотип АаВb; в потомстве F2 наблюдалось расщепление 9:7, характерное для комплементарного наследования.
Задача 5
У лука ген R определяет красную окраску чешуй, а ген r – желтую. Любая окраска проявляется только при наличии в генотипе доминантного гена С, при его отсутствии чешуи имеют белую окраску. Определить генотипы исходных форм луковиц с белыми и красными чешуями, если все гибридные луковицы имели красную окраску чешуй.
Решение:
R — ген, определяющий красную окраску чешуи лука
r — ген, определяющий жёлтую окраску чешуи лука
C — ген, способствующий проявлению окраски
c — не влияет на окраску
1. Белая окраска луковиц может определяться генотипами ссRR, ccRr или ccrr, а красная — CcRR, CcRr или CCRR. Известно, что при скрещивании луковиц с белыми и красными чешуями получено потомство всё с красными луковицами, значит, в его генотипе присутствуют оба доминантных гена R и C.
2. Все потомство единообразно, следовательно, скрещивались гомозиготные особи. 3. Один родитель имел неокрашенные чешуи, значит, он мог нести оба рецессивных гена (c и r); другой родитель имел красную окраску чешуи, значит, он мог нести оба доминантных гена (C и R).
4. Этим условиям удовлетворяет только тот вариант, когда у одного родителя генотип ссrr, а у другого – CCRR.
Ответ:
Генотип исходных растений с белыми чешуями луковиц – ссrr, с красными чешуями луковиц – CCRR.

На этом уроке подробно рассмотрены примеры решения задач на аллельное и неаллельное взаимодействие генов: кодоминирование, комплементарность, доминантный эпистаз, полимерию. Также этот урок включает пример решения задачи на плейотропное действие гена
В данный момент вы не можете посмотреть или раздать видеоурок ученикам
Чтобы получить доступ к этому и другим видеоурокам комплекта, вам нужно добавить его в личный кабинет, приобрев в каталоге.
Получите невероятные возможности



Конспект урока "Решение генетических задач. Взаимодействие генов"
Аллельное взаимодействие генов.
Кодоминирование.
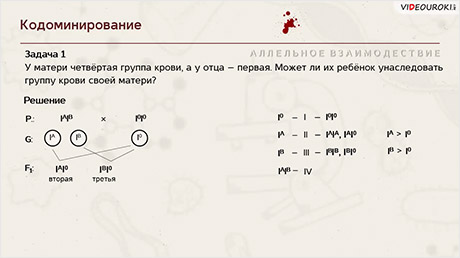
Задача 1. У матери четвёртая группа крови, а у отца – первая. Может ли их ребёнок унаследовать группу крови своей матери?
Решение: вспомним обозначения и комбинации генов, определяющие фенотипические признаки при наследовании групп крови у человека.

Возвращаемся к задаче.
Записываем генотипы родительских форм. У матери – I A I B , у отца – I 0 I 0 .
Определяем гаметы, которые дают родительские организмы. Поскольку от гомогаметного отца ребёнок не может получить ни ген I A , ни ген I B , то у него не может быть четвёртой группы крови. У половины потомков этих родителей может быть только вторая группа крови, а у другой половины – только третья.
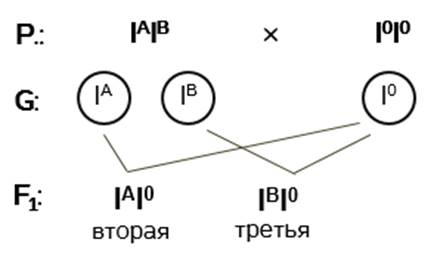
Ответ: нет, не может.
Задача 2. В родильном доме перепутали двух малышей. Первая пара родителей имеет I и II группы крови, вторая пара – II и IV. У одного из детей – II группа крови, а у второго – I группа. Помогите медперсоналу определить родителей обоих детей.
Решение: рассмотрим варианты генотипов первого ребёнка со второй группой крови. Это могут быть I A I A или I A I 0 . Анализируем генотипы родителей. И делаем вывод, что родителями ребёнка может быть как одна пара, так и другая.

Со вторым ребёнком всё значительно проще. Он гомогаметен, поскольку имеет первую группу крови. Значит, у обоих его родителей в генотипе должен быть ген I 0 . Такой вариант возможен только с первой парой родителей.
Ответ: ребёнок со второй группой крови – наследник второй пары родителей; ребёнок с первой группой крови рождён первой парой родителей.
Неаллельное взаимодействие генов.
Комплементарность (дополнительное взаимодействие).
Задача 3. У душистого горошка окраска цветков проявляется только при наличии двух доминантных генов: А и В. Если в генотипе имеется только один доминантный ген, то окраска не развивается. Какое потомство в первом и втором поколении получится от скрещивания растений с генотипами ААbb и ааВВ?
Решение: из условия задачи видно, что гены А и B отвечают за образование пигмента и его проявление. Но это происходит только у тех особей, в генотипе которых они встречаются вместе. В остальных случаях окраска не проявляется и цветки остаются белыми.
Записываем данные нам генотипы родительских форм. По фенотипу это два белых растения. Поскольку они гомозиготы, то образуют по одному типу гамет: Ab и aB.
В первом поколении получаем единообразие гибридов: 100% красноцветковых дигетерозигот.

Скрещиваем гибридов первого поколения между собой.
Дигетерозиготные особи дают по четыре типа гамет.
Строим решётку Пеннета.
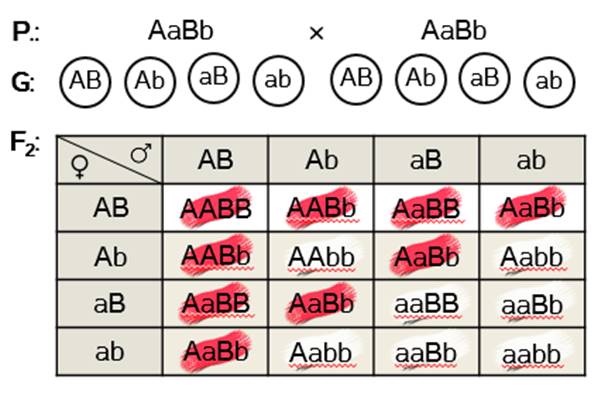
Во втором поколении гибридов получаем два варианта потомков по фенотипу – крас-ные и белые. Девять красных и семь белых. Девять к семи – это одно из соотношений расщепления, характерное для комплементарного взаимодействия генов.
Ответ: в первом поколении гибридов все особи с красными цветками дигетерозиготные. Во втором – наблюдаем расщепление: девять красноцветковых растений к семи белоцветковым.
Задача 4. При скрещивании растений одного из сортов тыквы с жёлтыми и белыми плодами всё потомство первого поколения имело белые плоды. При скрещивании гибридов F1 между собой во втором поколении было получено: 204 растения с белыми плодами, 53 – с жёлтыми и 17 – с зелёными. Определите генотип родителей и тип наследования.
Решение: определяем тип наследования. Для этого составляем соотношение расщепления.

Получаем 12:3:1. Такая пропорция говорит нам о том, что это эпистаз.
Вводим буквенные обозначения генов и записываем признаки, которые эти гены определяют. Пускай А – ген, определяющий жёлтую окраску, а – зелёную. Ген B подавляет образование пигмента, а b – не оказывает влияния на окраску.
Расщепление во втором поколении 12:3:1 означает, что родительские формы этих гибридов были дигетерозиготными, поскольку дали четыре типа гамет. А так как в первом поколении гибридов наблюдалось их единообразие, значит родительские формы гибридов первого поколения были гомозиготами по двум признакам. Одни из аллелей этих гомозигот доминантные, а другие – рецессивные.
Записываем генотипы родительских форм гибридов первого поколения: AAbb и aaBB. Проверяем соответствие записанных генотипов с фенотипами, данными по условию задачи. Одни родительские формы жёлтые, другие – белые.
Получаемое первое поколение гибридов – белые дигетерозиготы.

Скрещиваем их между собой.
Получаем заданное расщепление по фенотипу у гибридов второго поколения.
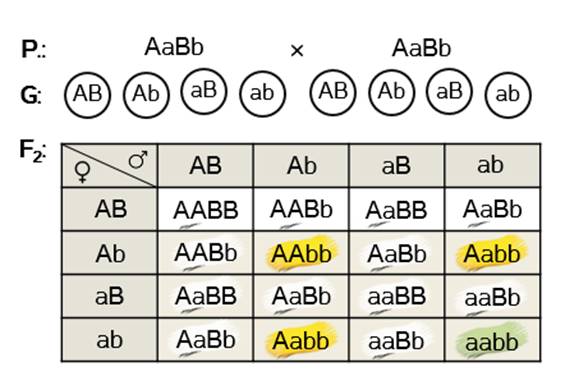
Записываем ответ задачи. Родители: доминантная гомозигота по первому признаку и рецессивная гомозигота по второму, и рецессивная гомозигота по первому признаку, доминантная гомозигота по второму. Тип наследования – доминантный эпистаз.
Задача 5. Рост человека определяется взаимодействием нескольких пар генов: А1a1A2a2А3a3. Люди с генотипом a1a1a2a2a3a3 имеют рост 150 см. С генотипом А1А1A2A2А3А3 – около 180 см (каждый доминантный ген добавляет к росту 5 см). Племя людей низкого роста порабощается ордой воинов с ростом в 180 см. Победители убивают мужчин и женятся на их женщинах. Какой рост будет у детей первого поколения от этих браков?
Решение: все гены отвечают за рост человека.
Записываем генотипы родителей. По условию – это доминантные и рецессивные гомозиготы.
Гомозиготы дают по одному типу гамет. В результате гибриды первого поколения будут гетерозиготами по трём парам генов.
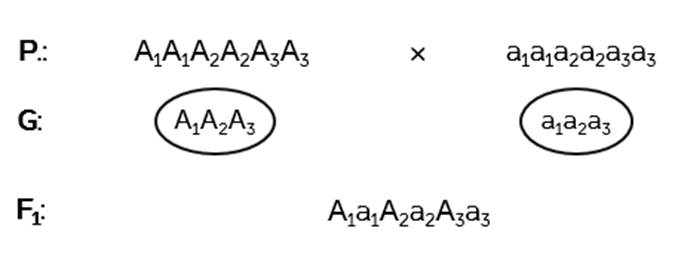
Поскольку каждый доминантный ген прибавляет к самому малому росту в 150 сантиметров по 5 см, а доминантных гена в данном случае три, значит потомки будут иметь рост 165 сантиметров.
Ответ: 165 см.
Плейотропия.
Задача 6. У мышей ген доминантной жёлтой пигментации шерсти обладает гомозиготным летальным действием. Его аллель определяет рецессивную чёрную пигментацию и обеспечивает жизнеспособность мышей. Скрещены две жёлтые особи. Какое расщепление по окраске шерсти ожидается у гибридов первого поколения?
Решение: вводим буквенные обозначения генов. Пускай ген А обуславливает жёлтую окраску шерсти, а ген а – чёрную.
При этом отмечаем, что в гомозиготном состоянии доминантные гены приводят к нежизнеспособности потомства.
Записываем генотипы родительских форм.
Жёлтые жизнеспособные мыши могут быть только гетерозиготами.
Записываем генотипы гибридов первого поколения.

Получаем четыре части гибридов, из которых одна часть доминантных гомозигот оказывается нежизнеспособной.
Ответ: живыми в потомстве окажутся две части мышей с жёлтой шерстью и одна часть – с чёрной.
Вопросы для повторения и задания
2. Чем определяется существующая специализация клеток?
3. Какие обязательные элементы входят в состав гена эукариотической клетки?
4. Приведите примеры взаимодействия генов.
Подумайте! Выполните!
1. Митохондрии содержат ДНК, гены которой кодируют синтез многих белков, необходимых для построения и функционирования этих органоидов. Подумайте, как будут наследоваться эти внеядерные гены.
2. Вспомните известные вам особенности развития человека. На каком этапе эмбриогенеза уже возникает чёткая дифференциация клеток?
Работа с компьютером
Обратитесь к электронному приложению. Изучите материал и выполните задания.
Взаимодействие неаллельных генов. Известно несколько видов взаимодействия неаллельных генов.
Комплементарное взаимодействие. Явление взаимодействия нескольких неаллельных генов, приводящее к развитию нового проявления признака, отсутствующего у родителей, называют комплементарным взаимодействием. Пример наследования окраски цветка у душистого горошка, приведённый в § 28, относится как раз к этому типу взаимодействия генов. Доминантные аллели двух генов (А и В) каждый в отдельности не могут обеспечить синтез пигмента. Антоциановый пигмент, вызывающий пурпурную окраску цветка, начинает синтезироваться только в том случае, когда в генотипе присутствуют доминантные аллели обоих генов (А_В_) (рис. 83).
Рис. 83. Наследование окраски венчика у душистого горошка
Рис. 84. Наследование формы гребня у кур
Известным примером комплементарного взаимодействия является наследование формы гребня у кур (рис. 84). Существует четыре формы гребня, формирование которых определяется взаимодействием двух неаллельных генов – А и В. При наличии в генотипе доминантных аллелей только гена А (А_bb) образуется розовидный гребень, наличие доминантных аллелей второго гена В (aaB_) обусловливает образование гороховидного гребня. Если в генотипе присутствуют доминантные аллели обоих генов (А_В_), образуется ореховидный гребень, а при отсутствии доминантных аллелей (aabb) развивается простой гребень.
Эпистаз. Взаимодействие неаллельных генов, при котором ген одной аллельной пары подавляет проявление гена другой аллельной пары, называют эпистазом. Гены, которые подавляют действие других генов, называют ингибиторами или супрессорами. Гены-ингибиторы могут быть как доминантными (I), так и рецессивными (i), поэтому различают доминантный и рецессивный эпистазы.
При доминантном эпистазе один доминантный ген (I) подавляет проявление другого неаллельного доминантного гена.
Возможны два варианта расщепления по фенотипу при доминантном эпистазе.
1. Гомозиготы по рецессивным аллелям (aaii) фенотипически не отличаются от организмов, имеющих в своём генотипе доминантные аллели гена-ингибитора. У тыквы окраска плода может быть жёлтой (А) и зелёной (а) (рис. 85). Проявление этой окраски может быть подавлено доминантным геном-ингибитором (I), в результате чего сформируются белые плоды (А_I_; aaI_).
В описанном и аналогичных случаях при расщеплении в F2 по генотипу 9:3:3:1 расщепление по фенотипу соответствует 12:3:1.
2. Гомозиготы по рецессивным аллелям (aaii) не отличаются по фенотипу от организмов с генотипами A_I_ и aaI_.
У кукурузы структурный ген А определяет окраску зерна: пурпурная (А) или белая (а). При наличии доминантного аллеля гена-ингибитора (I) пигмент не синтезируется.
Рис. 85. Наследование окраски плода у тыквы
В F2 у 9/16 растений (A_I_) пигмент не синтезируется, потому что в генотипе присутствует доминантный аллель гена-ингибитора (I). У 3/16 растений (aaI_) окраска зерна белая, так как в их генотипе нет доминантного аллеля А, отвечающего за синтез пигмента, и, кроме того, присутствует доминантный аллель гена-ингибитора. У 1/16 растений (aaii) зёрна тоже белые, потому что в их генотипе нет доминантного аллеля А, отвечающего за синтез пурпурного пигмента. Только у 3/16 растений, имеющих генотип A_ii, формируются окрашенные (пурпурные) зёрна, так как при наличии доминантного аллеля А в их генотипе отсутствует доминантный аллель гена ингибитора.
В этом и других аналогичных примерах расщепление по фенотипу в F2 13:3. (Обратите внимание, что по генотипу расщепление всё равно остаётся прежним – 9:3:3:1, соответствующим расщеплению в дигибридном скрещивании.)
При рецессивном эпистазе рецессивный аллель гена – ингибитора в гомозиготном состоянии подавляет проявление неаллельного доминантного гена.
У льна ген В определяет пигментацию венчика: аллель В – голубой венчик, аллель b – розовый. Окраска развивается только при наличии в генотипе доминантного аллеля другого неаллельного гена – I. Присутствие в генотипе двух рецессивных аллелей ii приводит к формированию неокрашенного (белого) венчика.
При рецессивном эпистазе в этом и других аналогичных случаях в F2 наблюдается расщепление по фенотипу 9:3:4.
Полимерное действие генов (полимерия). Ещё одним вариантом взаимодействия неаллельных генов является полимерия. При таком взаимодействии степень выраженности признака зависит от числа доминантных аллелей этих генов в генотипе: чем больше в сумме доминантных аллелей, тем сильнее выражен признак. Примером такого полимерного взаимодействия является наследование окраски зёрен у пшеницы (рис. 86). Растения с генотипом А1А1А2А2 имеют тёмно-красные зёрна, растения a1a1a2a2 – белые зёрна, а растения с одним, двумя или тремя доминантными аллелями – разную степень окраски: от розовой до красной. Такую полимерию называют накопительной или кумулятивной.
Однако существуют варианты и некумулятивной полимерии. Например, наследование формы стручка у пастушьей сумки определяется двумя неаллельными генами – А1 и А2. При наличии в генотипе хотя бы одного доминантного аллеля формируется треугольная форма стручка, при отсутствии доминантных аллелей (a1a1a2a2) стручок имеет овальную форму. В этом случае расщепление во втором поколении по фенотипу будет 15:1.
Рис. 86. Наследование окраски зёрен пшеницы
29. Генетика пола
Вспомните!
Каково соотношение мужчин и женщин в человеческой популяции?
Что вам известно об определении пола из предыдущих курсов биологии?
Какие организмы называют гермафродитными?
Проблема взаимоотношения полов, морфологические и физиологические отличия мужчин и женщин, их особенности темперамента и поведения всегда интересовали человечество. От чего зависит пол новорождённого младенца? Нельзя ли предсказать или предопределить рождение ребёнка определённого пола? Почему у раздельнополых видов, к которым относится человек, численность мужских и женских особей, как правило, примерно одинакова? Наконец, почему у одной и той же пары родителей рождаются потомки разного пола?

Моногибридное скрещивание. Мендель начал свои исследования закономерностей наследования с моногибридного скрещивания. Он выбрал две чистые линии растений гороха, которые отличались только по одному признаку: у одних окраска горошин была всегда жёлтая, а у других – всегда зелёная (при условии самоопыления). Если пользоваться современной терминологией, то можно сказать, что клетки растений гороха одного сорта содержат по два гена, кодирующих только жёлтую окраску, а другого сорта – по два гена, кодирующих только зелёную окраску семян. Гены, ответственные за проявление одного признака (например, формы или цвета семян), получили название аллельных генов. Если организм содержит два одинаковых аллельных гена (например, оба гена зеленого цвета семян или, наоборот, оба гена жёлтого цвета), то такие организмы называют гомозиготными. Если же аллельные гены различны (например, если один из них определяет жёлтую окраску семян, а другой – зелёную), то такие организмы называют гетерозиготными. Чистые линии образованы только гомозиготными растениями, поэтому при самоопылении они всегда воспроизводят один вариант проявления признака. В опытах Менделя, например, это был один из двух возможных цветов семян гороха – или всегда жёлтый, или всегда зелёный.
Правило единообразия гибридов первого поколения. Г. Мендель начал свои исследования со скрещивания растений гороха, исходно отличающихся только цветом горошин (жёлтым или зелёным). В первом поколении семена у всех растений оказались исключительно жёлтыми. Когда Г. Мендель повторил свои опыты по моногибридному скрещиванию, но использовал в них растения, отличающиеся друг от друга по другому признаку, по форме семян (гладкие или морщинистые), то все гибридные растения первого поколения имели гладкие семена. Проявляющиеся у гибридов признаки (желтизну или гладкость семян) Мендель назвал доминантными, а подавляемые признаки (зелёный цвет или морщинистую форму семян) – рецессивными. Доминантный признак принято обозначать прописными латинскими буквами (А, Б, С), а рецессивные – строчными (а, b, с).
На основе полученных в своих экспериментах данных Г. Мендель сформулировал правило единообразия гибридов первого поколения: при скрещивании двух гомозиготных организмов, отличающихся друг от друга одним признаком, все гибриды первого поколения будут иметь признак одного из родителей, и поколение по данному признаку будет единообразным.
Правило расщепления. Г. Мендель продолжил свои опыты, вырастив растения гороха из семян, полученных в первом поколении. Затем он скрестил эти растения и обнаружил, что у растений второго поколения большинство горошин, а именно 3 /4, были жёлтыми, а меньшая часть, а именно 1 /4, – зелёными. Конечно, Г. Мендель подсчитывал число жёлтых и зелёных горошин в потомстве от многих пар скрещиваемых растений гороха, чтобы добиться статистической надёжности полученного результата.
Явление, при котором скрещивание приводит к образованию части потомства с доминантным, а части – с рецессивным признаком, получило название расщепления.
Затем Г. Мендель подтвердил характер расщепления в опытах с другими признаками растений гороха и обосновал правило расщепления: при скрещивании двух потомков (гибридов) первого поколения между собой во втором поколении наблюдается расщепление и снова появляются особи с рецессивными признаками; эти особи составляют 1 /4 часть от всего числа потомков второго поколения.

Рис. 57. Цитологические основы моногибридного скрещивания
Цитологические основы закономерностей наследования при моногибридном скрещивании. Как можно схематически представить себе закономерности наследования признаков, открытые Г. Менделем, используя современные понятия?
Символ обозначает женскую особь, символ – мужскую, X – скрещивание, Р – родительское поколение, F1 – первое поколение потомков, F2 – второе поколение потомков, А – доминантный ген, отвечающий за жёлтый цвет горошин, а – рецессивный ген, отвечающий за зелёный цвет семян гороха (рис. 57).
В результате мейоза в гаметах родительских особей будут присутствовать по одному гену, отвечающему за наследование цвета семян: в случае женской гаметы – А, в случае мужской – а. В первом поколении (F1) соматические клетки будут гетерозиготными (Аа), поэтому половина гамет гибридов первого поколения будет содержать ген А, а другая половина – а. В результате случайных комбинаций гамет во втором поколении (F2) возникнут следующие комбинации: АА, Аа, аА, аа. Растения с тремя первыми комбинациями генов будут иметь жёлтые семена, а с четвёртой – зелёные.
Аллельные гены. Гомозиготы. Гетерозиготы. Доминантные и рецессивные гены. Правило единообразия. Правило расщепления. Закон чистоты гамет.
1. Почему Г. Мендель опылял растения гороха искусственно?
2. Какие организмы называются гомозиготными по какому-либо признаку?
§ 40. Множественные аллели. Анализирующее скрещивание
1. Какое доминирование называется неполным?
2. Что такое фенотип; генотип?
Неполное доминирование. Иногда один аллельный ген не до конца подавляет действие второго. При этом возникают промежуточные признаки, и признак у гомозиготной по доминантному гену особи будет не таким, как у гетерозиготной особи. Это явление получило название неполного доминирования. Поясним его на примере.

Рис. 58. Схема наследования признака в случае неполного доминирования
При скрещивании растения ночной красавицы, имеющего пурпурные цветки (АА), с растением, имеющим белые цветки (аа), все растения – гибриды первого поколения будут иметь розовые цветки (рис. 58). При скрещивании двух особей ночной красавицы из первого поколения во втором поколении происходит расщепление, но не 3: 1 (как при полном доминировании), а в другом соотношении – 1:2:1, т. е. 1 /4 растений с белыми цветками (аа), 2 /4 – с розовыми (Аа) и 1 /4 – с пурпурными (АА) (см. рис. 58).
Явление неполного доминирования встречается часто, например при наследовании цвета шерсти крупного рогатого скота, строения перьев птиц. Встречается это явление и у человека при наследовании брахидактилии – укорочения фаланг пальцев. У здорового человека имеется два доминантных гена ВВ, и развитие скелета происходит нормально. У гетерозигот (Вb) наблюдается укорочение фаланг пальцев, а у рецессивных гомозигот (bb) проявляются множественные нарушения в развитии скелета, и такие люди погибают ещё в раннем детстве. Таким образом, ген В не полностью подавляет действие гена b, и у гетерозигот вследствие этого развивается брахидактилия.
Сверхдоминирование. Изредка при взаимодействии двух аллельных генов наблюдается явление сверхдоминирования. При сверхдоминировании доминантный признак сильнее проявляется у гетерозигот (Аа), чем у доминантных гомозигот (АА). В частности, были выведены сорта томатов с набором аллельных генов Аа, который определяет их более высокую урожайность по сравнению с особями, имеющими набор АА.
До сих пор мы рассматривали случай наследования одного признака, определяемого одним геном. Но любой организм имеет громадное количество признаков, причём это не только внешние, видимые особенности, но и признаки биохимические (строение молекул, активность ферментов, концентрация веществ в тканях, крови и т. д.), анатомические (форма и размер внутренних органов) и т. п. Любой признак, каким бы простым он ни казался, определяется множеством физиологических и биохимических процессов, каждый из которых, в свою очередь, зависит от действия ферментов.
Совокупность всех внешних и внутренних признаков и свойств организма называют фенотипом.
Совокупность всех генов организма называют генотипом.
Фенотипические признаки, определяемые теми или иными генами, в различных условиях существования проявляются по-разному.
По фенотипу не всегда можно определить, какие гены содержит данная особь. Например, у растения гороха, имеющего жёлтые семена, генотип может быть как АА, так и Аа. А вот рецессивный признак – зелёный цвет семян – проявляется только у гомозиготных растений с генотипом аа. Иными словами, всегда можно определить генотип особи с рецессивным признаком.
Анализирующее скрещивание. Для установления генотипа особей, которые не различаются по фенотипу, используют так называемое анализирующее скрещивание. При этом особь, генотип которой нужно установить, скрещивают с особью, гомозиготной по рецессивному гену (аа). Например, для того чтобы выяснить, какие из растений гороха с жёлтыми семенами имеют генотип АА, а какие – Аа, их следует скрестить с растением с зелёными семенами (аа). Если потомство будет иметь только жёлтые горошины, значит, исследуемое растение было доминантной гомозиготой (АА). Если же в потомстве наблюдается расщепление на растения с жёлтыми и зелёными горошинами в соотношении 1:1, то изучаемое растение было гетерозиготно (Аа).
Генофонд. Итак, благодаря множественному аллелизму каждый вид имеет целый набор аллельных генов, хотя каждая особь благодаря диплоидности содержит в каждой аллели только два гена. Совокупность всех вариантов всех генов, входящих в состав генотипов особей какого-либо вида, получила название генофонда вида. Можно также говорить о генофонде популяции или иной группы особей вида.
Множественный аллелизм. Кодоминирование. Неполное доминирование. Сверхдоминирование. Фенотип. Генотип. Анализирующее скрещивание. Генофонд вида.
1. Что такое множественный аллелизм?
2. Какие виды доминирования вам известны?
3. Какое практическое значение может иметь изучение видов доминирования?
4. В чём отличие двух понятий: фенотип и генотип?
5. Какой метод используется для установления генотипа особей, не различающихся по фенотипу?
6. Как наследуется брахидактилия?
§ 41. Дигибридное скрещивание. Закон независимого наследования признаков
1. Какое скрещивание называется моногибридным?
2. Сильно ли различается набор генов в клетках корня и клетках листа одной и той же особи клёна?
Дигибридное скрещивание. В природе не встречаются две абсолютно одинаковые особи какого-либо вида живых существ – все организмы отличаются друг от друга по многим признакам. Исключение могут представлять растения, развивающиеся в результате самоопыления (например, пшеница, горох, картофель и др.), а также однояйцевые близнецы животных и человека, хотя говорить об абсолютной схожести таких особей также нельзя.
Скрещивание особей, у которых учитывают отличия друг от друга по двум признакам, называется дигибридным, если по трём – то тригибридным и т. д. В общем случае скрещивание особей, отличающихся по многим признакам, называется полигибридным.
Закон независимого наследования признаков. Г. Мендель приступил к изучению результатов дигибридного скрещивания после того, как установил закономерности моногибридного скрещивания. Для этого он исследовал характер расщепления при скрещивании двух чистых линий гороха, различающихся по двум признакам: цвету семян (жёлтые или зелёные) и их форме (гладкие или морщинистые). При таком скрещивании признаки определяются различными парами генов: одна пара генов отвечает за цвет семян, другая – за их форму. При этом жёлтая окраска горошин (А) доминирует над зелёной (а), а их гладкая форма (B) – над морщинистой (b).
По закону единообразия гибридов первого поколения семена гороха в поколении F1 были жёлтыми и гладкими. Для того чтобы было легче понять, как будет проходить комбинация признаков при скрещивании двух гибридов из первого поколения, американский исследователь Реджинальд Пеннет предложил заносить результаты опыта в таблицу, которую назвали решёткой Пеннета (рис. 59). В результате слияния четырёх видов гамет, возникающих у растений из F1 (AB, Аb, аВ, аb), во втором поколении (F2) возникает 9 различных генотипов. Но эти девять генотипов проявляются в виде четырёх фенотипов: жёлтые – гладкие, жёлтые – морщинистые, зелёные – гладкие и зелёные – морщинистые, причём соотношение фенотипов будет 9:3:3:1. Если же подсчитать, каким будет соотношение между жёлтыми и зелёными горошинами, то оно будет равняться 3:1. Такое же соотношение будет и между гладкими и морщинистыми семенами. То есть выполняется правило расщепления. Отсюда следует, что при дигибридном скрещивании гены и признаки, за которые эти гены отвечают, наследуются независимо друг от друга. Этот факт получил название закона независимого наследования признаков. Закон справедлив в тех случаях, когда гены рассматриваемых признаков располагаются в разных негомологичных хромосомах.
Решётка Пеннета. Закон независимого наследования признаков.
1. Какое скрещивание называется дигибридным?
2. В чём сущность закона независимого наследования признаков?
3. В каком случае закон независимого наследования признаков несправедлив?
4. Сколько фенотипов гороха наблюдал Г. Мендель во втором поколении при дигибридном скрещивании гороха?

Рис. 59. Схема наследования признаков при дигибридном скрещивании
§ 42. Хромосомная теория наследственности
1. Что такое хромосомы? Какова их роль?
2. Знал ли Г. Мендель о существовании хромосом?
3. Можно ли назвать гены, определяющие цвет семян гороха, и гены, определяющие цвет венчика цветков гороха, аллельными?
В 1902 г. американец Уильям Сеттон предположил, что элементы наследственности, которые мы сегодня называем генами, могут располагаться в хромосомах. Но вместе с тем стали накапливаться сведения о том, что в некоторых случаях расщепления по правилам Менделя не происходит. Например, у душистого горошка два признака – форма пыльцы и окраска цветков – не дают независимого расщепления в потомстве в соотношении 3:1, и потомки остаются похожими на родительские особи. При последующем анализе оказалось, что гены этих двух признаков лежат в одной хромосоме.
Действительно, генов, кодирующих различные признаки, у любого организма очень много. Так, по приблизительным подсчётам, у человека 30–40 тыс. генов, а видов хромосом всего 23. Все это огромное количество генов размещается в этих хромосомах.
Каковы же принципы наследования генов, расположенных на одной хромосоме? Современная хромосомная теория наследственности создана выдающимся американским генетиком Томасом Морганом (1866–1945).
Первое положение этой теории гласит: ген представляет собой участок хромосомы. Хромосомы, таким образом, представляют собой группы сцепления генов.
Второе положение утверждает, что аллельные гены (гены, отвечающие за один признак) расположены в строго определённых местах (локусах) гомологических хромосом.
Наконец, согласно третьему положению, гены располагаются в хромосомах линейно, т. е. друг за другом.
Основным объектом, с которым работали Морган и его ученики, была плодовая мушка дрозофила, имеющая диплоидный набор из 8 хромосом. Эксперименты показали, что гены, находящиеся в одной хромосоме, при мейозе попадают в одну гамету, т. е. наследуются сцепленно. Это явление получило название закона Моргана.
Кроссинговер. Однако в тех же опытах было описано и отклонение от этого закона. Среди гибридов второго поколения обязательно было малое число особей с перекомбинацией тех признаков, гены которых лежат в одной хромосоме. Как это можно объяснить?
Для этого необходимо вспомнить ход мейоза, а именно то, что в профазу первого мейотического деления гомологичные хромосомы конъюгируют и могут обмениваться гомологичными участками. Этот процесс, как мы помним, называется кроссинговер; он очень важен для повышения разнообразия потомков. Кроссинговер также был открыт Т. Морганом и его сотрудниками, поэтому хромосомную теорию можно дополнить ещё одним, четвёртым, положением: в процессе образования гамет между гомологичными хромосомами происходит конъюгация, в результате которой они могут обмениваться аллельными генами, т. е. может происходить кроссинговер.
Таким образом, при кроссинговере происходит нарушение закона Моргана, и гены одной хромосомы не наследуются сцепленно, так как часть из них заменяется на аллельные гены гомологичной хромосомы. Иными словами, сцепление генов является неполным.
Хромосомные карты. Явление кроссинговера помогло учёным установить расположение каждого гена в хромосоме, создать генетические карты хромосом. Морган и его сотрудники сделали вывод о том, что чем дальше друг от друга расположены на хромосоме два гена, тем чаще они будут расходиться в разные хромосомы в процессе кроссинговера. Если два гена А и D расположены на противоположных концах одной хромосомы (рис. 60), то они разойдутся в разные хромосомы при любом варианте кроссинговера. Но если речь идёт о двух генах Б и С, расположенных в хромосоме рядом, то они могут разойтись по разным хромосомам только в том случае, если точка перекреста окажется на одном-единственном участке между этими двумя генами. Вероятность такого события не очень велика. Таким образом, вероятность расхождения двух генов по разным хромосомам в процессе кроссинговера зависит от расстояния между ними в хромосоме. Следовательно, подсчитав частоту кроссинговера между какими-либо двумя генами одной хромосомы, отвечающими за различные признаки, можно точно определить расстояние между этими генами, а значит, и начать построение генетической карты, которая представляет собой схему взаимного расположения генов, составляющих одну хромосому.

Рис. 60. Возможные варианты кроссинговера, происходящие между гомологичными хромосомами (вероятность расхождения двух генов в различные хромосомы тем выше, чем дальше друг от друга они расположены)
Хромосомная теория наследственности. Закон Моргана. Кроссинговер. Генетические карты.
1. Что представляет собой ген?
2. В каких случаях справедливо правило независимого наследования признаков?
Читайте также: