
Какие признаки положены в основу классификации механических тканей
Обновлено: 04.07.2024
- Механи́ческая ткань — вид ткани в растительном организме, волокна из живых и мёртвых клеток с сильно утолщённой клеточной стенкой, придающие механическую прочность организму. Возникает из верхушечной меристемы, а также в результате деятельности прокамбия и камбия.
Степень развития механических тканей во многом зависит от условий, они мало присутствуют у растений влажных лесов, у многих прибрежных растений, но зато хорошо развиты у большинства растений засушливых местообитаний.
Механические ткани присутствуют во всех органах растения, но наиболее они развиты по периферии стебля и в центральной части корня.
Выделяют следующие типы механических тканей:
* колленхима — эластичная опорная ткань первичной коры молодых стеблей двудольных растений, а также листьев. Состоит из живых клеток с неравномерно утолщёнными неодеревеневшими первичными оболочками, вытянутыми вдоль оси органа. Создаёт опору растению.
* склеренхима — прочная ткань из быстро отмирающих клеток с одревесневшими и равномерно утолщёнными оболочками. Обеспечивает прочность органов и всего тела растений. Различают два типа склеренхимных клеток:
* волокна — длинные тонкие клетки, обычно собранные в тяжи или пучки (например, лубяные или древесинные волокна).
Связанные понятия
Колленхи́ма (др.-греч. κόλλᾰ — клей) — одна из первичных механических тканей растений, располагающаяся в первичной коре стеблей и листьях в основном у двудольных растений. Механическая функция колленхимы основана на осмотических явлениях.
Кора́ (лат. cortex) — общее название совокупности тканей, расположенных снаружи от камбия. Имеется как в стеблях, так и в корнях, состоит из тканей различного строения и происхождения. В общей сложности в состав коры можно включить следующие растительные ткани: корка (ритидом), перидерма (пробка (или феллема), феллоген, феллодерма), первичная кора, перицикл, вторичная кора (вторичная флоэма).
Ко́рень (лат. radix) — осевой, обычно подземный вегетативный орган высших сосудистых растений, обладающий неограниченным ростом в длину и положительным геотропизмом. Корень осуществляет закрепление растения в почве и обеспечивает поглощение и проведение воды с растворёнными минеральными веществами к стеблю и листьям.
Трагака́нт — высохшая на воздухе камедь (гуммитрагакант, лат. Gummi Tragacanthae), которая вытекает из трещин или надрезов стеблей или ветвей некоторых растений рода Астрагал (Astragalus) семейства Бобовые (Fabaceae) в результате перерождения клеточных стенок паренхимы сердцевины и сердцевинных лучей.
Упоминания в литературе
Механические ткани – это опорные ткани. Они придают растению прочность благодаря утолщениям их клеточных стенок и соответствующему распределению в органе растения. Различают два типа механической ткани – колленхима и склеренхима.
В поперечном сечении форма стеблей также очень разнообразна. Поперечные срезы стеблей помогают различить круглые стебли (такие стебли встречаются у ромашки, большого количества злаков, дремы). Округлые стебли встречаются у чеснока, сплюснутые – у мятлика, существуют также трехгранные, четырехгранные, многогранные, гранистые, бороздчато-гранистые, крылато-гранистые стебли и очень много более сложных форм. При поперечном разрезе стебля можно увидеть воздушные полости, затем элементы проводящей и механической тканей . Полость называется совершенной в том случае, когда у полых стеблей их центральная полость имеет на поперечном срезе совершенно ровные края, например в соломине злаков. Если центральная полость ограничена неровной линией, то она называется несовершенной.
Калий существенно влияет на величину урожая картофеля и его качество (особенно крахмалистость), повышает устойчивость растений к болезням. Калий играет исключительную роль в водном режиме растений. При калийном голодании картофеля происходят нарушения в росте и развитии растения. Механические ткани и корневая система развиваются слабее. Клубни при недостатке калия бывают мелкими, приобретают несколько удлиненную форму и плохо хранятся в зимний период. Калийные удобрения, содержащие много хлора, уменьшают размер крахмального зерна.
Лист – боковой орган растения, который обеспечивает функции фотосинтеза, транспирации и газообмена. Лист состоит из листовой пластинки и черешка. Листовые пластинки могут быть цельными или рассеченными. Листья различаются по форме листовой пластинки (округлые, ланцетовидные, овальные, игольчатые и т.д.) и по форме края листовой пластинки (цельнокрайние, зубчатые, выемчатые). Расположение листьев на стебле может быть очередным, супротивным, мутовчатым. Различают простые и сложные листья. У сложных имеется несколько листовых пластин, прикрепленных к основному черешку черешочками. Лист снаружи и снизу покрыт эпидермой, между слоями которой находится мякоть листа – мезофил, содержащий хлорофиллоносную ткань, а также сосудисто – волокнистые пучки и механическую ткань (рисунок 9).
Связанные понятия (продолжение)
Ксиле́ма (от греч. ξύλον — древеси́на) — основная водопроводящая ткань наземных сосудистых растений; один из двух подтипов проводящей ткани растений, наряду с флоэмой — лубом.
Абсорбцио́нные тка́ни — ткани растений, поглощающие жидкие или газообразные вещества из внешней среды.
Проводящая ткань — вид тканей растений, служащих для передвижения по организму растворённых питательных веществ. У многих высших растений она представлена проводящими элементами (сосудами и ситовидными трубками). В стенках проводящих элементов есть поры и сквозные отверстия, облегчающие передвижение веществ от клетки к клетке.
Флоэ́ма (от греч. φλοῦς — кора) — то же, что и луб — проводящая ткань сосудистых растений, по которой происходит транспорт продуктов фотосинтеза к частям растения, в которых он не происходит: подземные части, конусы нарастания, цветки, плоды и др. Вместе с ксилемой (древесиной), обеспечивающей транспорт воды и минеральных солей, образует проводящие пучки.
Кутикула растений (от лат. cuticula — корка, надкожица) — защитный слой на поверхности растений, образуется с помощью эпидермальных клеток листьев, молодых побегов и других воздушных органов растений, не покрытых перидермой. Кутикула обычно толще на верхней стороне листа, хотя, вопреки распространенному мнению, толще на нижней стороне в ксерофитных растениях сухих климатических зон (по сравнению с мезофитными растениями влажных районов). Слой состоит из воскоподобного вещества кутина, покрывающий.
Эпиде́рма (эпиде́рмис, ко́жица) — внешняя первичная покровная ткань растений, обычно однослойная, покрывающая молодые стебли и остальные наземные органы (листья, лепестки, плоды и др.). Представляет собой наружный слой клеток, образующийся из протодермы конуса нарастания.
Периде́рма (от греч. περι — около и греч. δερμα — кожа) — комплекс тканей, состоящий из феллогена и его производных — феллодермы (откладывается внутрь) и феллемы, или пробки (вторичной покровной ткани, откладывается наружу). Перидерма является одним из чётко выраженных конструктивных элементов строения стебля высших растений, которые невозможно отнести ни к тканям, ни к органам. Такие элементы называются анатомо-топографическими зонами.
Камбиформ — растительная ткань, состоящая из клеток, видом весьма похожих на камбий (отсюда и название), но утративших способность делиться, — следовательно, камбиформ ткань постоянная. Он входит в состав луба (флоэмы) замкнутых сосудистых пучков однодольных, где, по мнению некоторых ученых, наряду с ситовидными трубками и клетками-спутниками (Geleitzellen), служит проводником пластических веществ.
Экзоде́рма — наружный слой клеток (реже — несколько слоёв), выделяемый в первичной коре корней некоторых растений. Экзодерма непосредственно подстилает ризодерму и характеризуется плотным расположением клеток (отсутствием межклетников) и наличием поясков Каспари, препятствующих апопластическому транспорту водных растворов в радиальном направлении. Образование поясков Каспари в экзодерме происходит в более старых частях корня, чем в эндодерме (за пределами зоны всасывания). Со временем стенки клеток.
Трахеи́ды — прозенхимные, мёртвые клетки ксилемы длиной в несколько миллиметров, шириной в десятые и сотые доли миллиметра, с утолщёнными одревесневшими оболочками, несущими поры (часто окаймлённые), через которые происходит фильтрация растворов из одной трахеиды в другую.
Хлоре́нхима, или хлорофиллоно́сная паренхи́ма, — ассимиляционная (то есть осуществляющая синтез молекулярных компонентов клетки) ткань сосудистых растений, состоящая из паренхимных клеток, вдоль тонких стенок которых одним слоем располагаются хлоропласты, не затеняя друг друга.
Расте́ния (лат. Plantae) — биологическое царство, одна из основных групп многоклеточных организмов, включающая в себя в том числе мхи, папоротники, хвощи, плауны, голосеменные и цветковые растения. Нередко к растениям относят также все водоросли или некоторые их группы. Растения (в первую очередь, цветковые) представлены многочисленными жизненными формами — среди них есть деревья, кустарники, травы и др.
Ха́ровые водоросли, или лучицы (лат. Charophyceae) — класс некогда обширной группы древних растений, которые объединяют в себе признаки водорослей и высших растений. Название происходит от др.-греч. χᾰρά — радость, красота. Всего известно не более 700 видов харовых.
То́поль чёрный, или Осоко́рь (лат. Pópulus nígra) — вид из рода Тополь семейства Ивовые. Медоносное, дубильное, эфиромасличное, красильное, лекарственное, древесинное, декоративное растение, культивируется в озеленении.
Анана́с (лат. Anánas) — род травянистых растений семейства Бромелиевые (Bromeliaceae), происходят из тропической Америки. Один из видов — Ананас хохлатый (Ananas comosus) — является важной плодовой культурой, широко возделывается в тропических странах по всему миру.
Гигрофи́ты (от др.-греч. ὑγρός — влажный + φυτόν — растение) — растения, обитающие в местах с высокой влажностью воздуха и (или) почвы. Категория гигрофитов выделяется на основе особенностей физиологии и морфологии растений; наиболее характерный признак гигрофитов — отсутствие приспособлений, ограничивающих расходование влаги.
Ка́мбий (от лат. cambium — обмен, смена) — образовательная ткань в стеблях и корнях преимущественно двудольных и голосеменных растений, дающая начало вторичным проводящим тканям и обеспечивающая их прирост в толщину. Сезонные изменения активности камбия обусловливают образование годичных колец древесины. Из клеток прокамбия или веретеновидных клеток камбия возникает камбиформ.
Хлопча́тник (лат. Gossypium) — род семейства Мальвовые (Malvaceae), включающий более 50 видов древесных и травянистых, многолетних, двулетних и однолетних растений, происходящих из тропических и субтропических районов Азии, Америки, Африки и Австралии. Культурные формы в промышленных масштабах выращивают по всему свету как прядильные растения. Является источником растительных волокон для текстильной промышленности — хлопка.
Эпибле́ма (греч. ἐπίβλημα – покрывало, покрытие от греч. ἐπί — на, над и греч. βλημα — бросаю, кладу) или ризоде́рма, — первичная покровная ткань молодых корней растений. Формирует корневые волоски.
Птерока́рпус мешкови́дный, или Птерока́рпус су́мчатый (лат. Pterocárpus marsupium) — крупное листопадное дерево, вид рода Птерокарпус (Pterocarpus) семейства Бобовые.
Микори́за (греч. μύκης — гриб и ρίζα — корень) (грибокорень) — симбиотическая ассоциация мицелия гриба с корнями высших растений.
Текстильные волокна — волокна, использующиеся в текстильной промышленности для изготовления текстильных материалов: ткани, нетканых материалов, трикотажных полотен, ниток, пряжи, а также искусственного меха.
Бамбу́ковое волокно́ — регенерированное целлюлозное волокно, изготовленное из стебля бамбука. Тонкостью и белизной напоминает вискозу, обладает более высокой прочностью.
Ствол — главный, осевой, радиально-симметричный, вегетативный орган растения, стебель древесных или древовидных (напр. пальмы) растений.
Склереи́ды — мертвые клетки механической ткани с толстыми одревесневшими оболочками (каменистые клетки).Клетки самой разнообразной формы, с равномерно утолщёнными слоистыми стенками, пронизанными простыми, нередко ветвистыми порами. Стенки склереид всегда сильно одревесневают, иногда пропитываются известью, кремнеземом и кутином. Живое содержимое, как правило, отмирает. Изредка, когда клетка-склереида сообщается с соседними живыми клетками, протопласт сохраняется, и в таких склереидах со временем.
Пыльца́, цветень — скопление пыльцевых зёрен семенных растений. Пыльцевое зерно представляет собой мужской гаметофит, развивающийся в микроспорангии из микроспоры и выполняющий функцию опыления, то есть оплодотворения женского гаметофита, находящегося в семязачатке.
Хло́пковое де́рево (лат. Ceiba pentandra) — тропическое дерево семейства мальвовых (ранее род Сейба относили к семейству Бомбаксовые). В естественных условиях произрастает в Мексике, Центральной Америке, Карибских островах, северной части Южной Америки и в тропиках западной Африки. Дерево являлось одним из священных символов мифологии майя. Также известно под именами Капок, Сейба пятитычинковая, Сумаума. Слово капок также применяется для названия волокна, получаемого из плодов этого дерева.
Печёночные мхи, или Печёночники, или Маршанциевые мхи (лат. Marchantiophyta ), — отдел растений со слабо развитой протонемой; распространены по всему земному шару, особенно в тропиках. Число видов — от шести до восьми тысяч.
Дерматоге́н (от греч. дермато и ген) — относительно однородный слой поверхностных клеток у растений, получивший своё название от того, что из него образуется впоследствии кожица (δέρμα — кожа). Расположен снаружи, имеет плотную структуру, относится к первичной образовательной ткани (меристема), расположен в растущих кончиках корней. Дерматоген имеет межклетники, его клетки обычно делятся так, что образующиеся перегородки имеют антиклинальное направление, то есть они растут перпендикулярно к поверхности.
Капиллиций, или волосяное сплетение (лат. Capillitium, восх. к лат. capillus — волос) представляет собой совокупность нитевидных волоконцев, которые образуют систему нитей в виде сетей и/или каркасов, которые содержатся в плодовых телах многих миксомицетов, а также у некоторых вздутых грибов (гастеромицетов). Капиллиций как орган способствует разрыхлению споровой массы и, благодаря гигроскопическим движениям, помогает рассеивать споры, и, таким образом, способствует процессу размножения. При анализе.
Сли́зевые ходы́ — длинные каналы в органах растений, замкнутые полости или отдельные клетки, заполненные слизистым веществом (слизью и камедью). Анатомически сходны со смоляными ходами. Возникают обычно схизогенно (путём расхождением клеток), иногда лизигенно (путём растворения клеток) и схизо-лизигенно (комбинированно). Ходы бывают простые и ветвистые.
Веламен (лат. velamen) — губчатая, обычно многослойная, гигроскопичная мёртвая ткань, покрывающая воздушные корни большинства эпифитных растений.
Сте́ла, или Стель, или Сте́ле, или Центра́льный цили́ндр, или Осево́й цилиндр — комплекс первичных тканей, лежащих внутри от первичной коры сосудистых растений.
Культу́ра тќаней — способ искусственного вегетативного размножения растений, а также способ культивирования in vitro органов или эксплантатов ткани животных.
Аэренхима (или эренхима) — воздухоносная ткань у растений, построенная из клеток, соединённых между собой так, что между ними остаются крупные заполненные воздухом пустоты (крупные межклетники).
Лист (множ. ли́стья, собир. листва́; лат. folium, греч. φύλλον) — в ботанике наружный орган растения, основными функциями которого является фотосинтез, газообмен и транспирация. Для этой цели лист, как правило, имеет пластинчатую структуру, чтобы дать клеткам, содержащим в хлоропластах специализированный пигмент хлорофилл, доступ к солнечному свету. Лист также является органом дыхания, испарения и гуттации (выделения капель воды) растения. Листья могут задерживать в себе воду и питательные вещества.
Симмондсия китайская (лат. Simmóndsia chinénsis), более известная под названиями Жожоба́ и Хохо́ба (исп. Jojoba) — вид ветвистых вечнозелёных кустарников, в диком виде встречающихся в североамериканских пустынях и чапарале. Это единственный вид рода Симмондсия (Simmondsia), который был выделен в отдельное монотипное семейство Симмондсиевые (Simmondsiaceae).
Корка, или ритидом (лат. rhytidoma) — наружная часть коры многолетних побегов и корней, состоит из омертвевших участков первичной коры и вторичной флоэмы. Эти две ткани разделяются перидермами, образуемыми неоднократно закладываемыми феллогенами. Периферические слои корки опадают, и старый слой феллогена отмирает. Вместо него дальше от центра закладывается новый слой, и, таким образом, формируется несколько перидерм. Отмирание тканей, располагающихся между перидермами, обусловлено газо- и водонепроницаемостью.
Макроцистис (лат. Macrocystis) — род бурых водорослей из семейства ламинариевых. Включает самые крупные из известных водорослей, превышающие длину 45 метров. Отдельные экземпляры живут до 8—10 лет, хотя обычно жизнь водоросли короче. Используются в аквакультуре. Обитают в Южном полушарии. Растут на скалистых и каменистых грунтах на глубине 20—30 м.
Основы учения о тканях были заложены еще в XVII веке, когда было установлено клеточное строение растений. Это открытие было сделано в 1665 г. англичанином Р. Гуком. Изучая под микроскопом срез бутылочной пробки, он обнаружил в нем систему замкнутых пузырьков, или ячеек. Впоследствии их стали называть клетками, хотя на самом деле эти ячейки представляли собой только оболочки клеток.
В 1807 г. Г. Ф. Линк (1767-1851) ввел понятие о паренхиматических и прозенхиматических клетках. Первую группу составляют клетки шаровидные, полиэдрические (изодиаметрические) или слегка призматические, вторую — более или менее длинные, с заостренными концами. Ткани, состоящие из этих клеток, стали называть соответственно паренхимой и прозенхимой .
Ботаники второй половины XIX века А. де Бари (1831-1888) и Ф. ван Тигем (1839-1914) называли тканью совокупность клеток одинакового строения, независимо от того, разбросаны они в теле растения или составляют компактные группы.
Классифицируя ткани, Ф. ван Тигем обращал внимание на наличие в клетках живого содержимого. По этому признаку он разделил ткани на живые и мертвые. К последним он отнес ткани, выполняющие в растении опорную роль и участвующие в проведении веществ. Условность такого разделения тканей очевидна: мертвые ткани на ранних стадиях развития состоят из живых клеток, а клетки живых тканей со временем могут терять содержимое и переходить в разряд мертвых.
Ю. Сакс (1832-1897) различал в растениях покровную , пучковую и основную ткани . Этой классификацией пользуются и в настоящее время. Неудобство её состоит в том, что в одну ткань часто попадают клетки, разные не только по строению, но и по выполняемым функциям. Так, пучковую ткань составляют клетки, проводящие воду и растворы органических веществ, а также клетки с толстыми одревесневшими оболочками, играющие механическую роль. Еще большее число функционально разных клеток объединяет основная ткань. Нередко к тому же участки этой ткани могут быть разрозненными, часто они находятся в разных частях растения и топографически не составляют непрерывную систему.
Пожалуй, наиболее удачное определение тканей, которым пользуются и в настоящее время, предложено в конце XIX века Ф. Габерландтом (1826-1878). Ткань — это устойчивый комплекс клеток, обладающих одним или несколькими сходными признаками: физиологическими, морфологическими, топографическими и общностью происхождения. Каждый из этих признаков может быть использован для классификации тканей, но, учитывая, что только согласованное функционирование разных тканей может обеспечить осуществление всех присущих любому организму жизненных процессов, наиболее целесообразна группировка тканей по анатомо-физиологическому принципу.
Ткани и локальные структуры, выполняющие одинаковые функции, Ф. Габерландт объединил в 9 систем.
Запасающая (эндосперм; перисперм; запасающая паренхима вегетативных органов; водозапасающие волоски);
Секреторная и выделительная (железистые волоски, внутренние железки, слизевые и смоляные ходы и клетки, масляные клетки, гидатоды, млечники);
Образовательная (меристема зародыша, конуса нарастания побега, кончика корня, прокамбий, камбий, феллоген, раневая меристема).
Строение тканей и их расположение в теле растения подчинено принципу максимальной физиологической активности. Наряду с главной, ткань может выполнять одну или несколько дополнительных функций. Это определяет морфологическую дифференциацию составляющих ее клеток.
Так, эпидерма, или кожица, не только защищает внутренние ткани от неблагоприятных факторов внешней среды, но участвует также в газообмене и транспирации, у многих растений она выполняет секреторную и выделительные функции благодаря железистым волоскам и накоплению в обычных клетках некоторых балластных продуктов метаболизма растений. Ксилема, или древесина, осуществляет восходящий ток воды с растворенными в ней минеральными веществами, обеспечивает механическую прочность растения, у многолетних растений некоторые ее клетки служат вместилищами запасных веществ. Этой полифункциональностью объясняется неоднородность ее строения. То же можно сказать о флоэме, или лубе. Такие ткани называют сложными в отличие от простых , состоящих из одинаковых по строению и функциям клеток. Простые ткани — это, например, колленхима, веламен, эндосперм.
Нередко внутри какой-либо одной специализированной ткани встречаются структуры, принадлежащие другой системе тканей. Так, в листьях некоторых растений между клетками ассимилирующей ткани могут развиваться склереиды — клетки с очень толстыми одревесневшими оболочками, принадлежащие к механической системе растений. Нередко в листьях встречаются вместилища эфирных масел, составляющие секреторную систему. Такие структуры, которые по строению и функциям отличаются от окружающей их ткани, называют идиобластами .
Наряду с анатомо-физиологической существует и онтогенетическая nклассификация тканей, основанная на их происхождении. По этой классификации ткани делят на первичные и вторичные.
Первичные ткани представляют собой непосредственные производные меристемы, находящейся на верхушке побега и в кончике корня (эпидерма, колленхима, склеренхима, ассимилирующая ткань, эпиблема), а также специализированной меристемы — прокамбия (первичная ксилема, первичная флоэма). Ко вторичным относят ткани, возникающие при утолщении стебля и корня. Это производные камбия (вторичная ксилема, вторичная флоэма), феллогена (пробка, феллодерма, чечевички), разные типы идиобластов. Вторичные ткани свойственны не всем растениям. Их нет у мхов, современных хвощей, плаунов, папоротников (за очень редким исключением), а из покрытосеменных — у большинства однодольных. Мощное развитие вторичных тканей, главным образом, древесины и луба, характерно для древесных растений.

Биология
Населяющие сушу растительные организмы постоянно подвергаются воздействию ветра, силы тяжести, снегопадов т. д. Кроме этого, они могут вытаптываться человеком или животными. Основная задача механической ткани растений заключается в противодействии нагрузкам на растяжение, сжатие и изгиб. Эти покровы являются своеобразным каркасом, пронизывающим все части растительного организма.
Классификация покровов
Растения имеют несколько типов тканей. Они различаются строением и выполняемыми функциями. Выделяют 6 типов покровов растительных организмов:
- Основной. Ткань представлена паренхимой и принимает активное участие во всех жизненно важных процессах — фотосинтезе, газообмене, создании запасов питательных веществ и т. д.
- Образовательный. Классифицируется на несколько типов покровов — вставочный, боковой, раневый, верхушечный. Основной задачей этой ткани является создание новых клеток и восстановление структур, получивших повреждения.
- Проводящий. Предназначена для доставки питательных элементов и воды к листочкам и стеблю.
- Выделительный. Основными функциями этого вида покрова растения является выведение лишней влаги, а также продуктов метаболизма.
- Покровный. Выделяют три типа этой ткани — пробка, эпидерма и корка. Они выполняют защитную функцию, и принимает участие в процессах газообмена.
- Механический.
Общая характеристика

Чтобы быстрее разобраться с функциями механического покрова растений, следует вспомнить различные неблагоприятные факторы внешней среды, воздействие которых они испытывают ежесекундно. Однако благодаря наличию в их структуре особой ткани, растительные организмы способны переносить сильный ветер, землетрясения, ливневые дожди и т. д.
Каждое растение в процессе эволюции приспособилось к среде своего обитания. Именно поэтому все виды тканей даже у растительных организмов одного вида концентрируются в их частях по-разному. Таким образом, функционал механической ткани обусловлен необходимостью защиты от негативного воздействия внешней среды.
Основная роль механической ткани заключается в сохранении целостности растения. В качестве примера можно привести деревья, которые под воздействием ветра гнуться, но не ломаются.
Строение механической ткани
Клетки этих структур устроены примерно одинаково. Они обладают толстыми стенками, что увеличивает устойчивость растения.
Клетки колленхимы
В основе клеточек колленхимы находятся два полисахарида: гемицеллюлаза и целлюлоза. Это один из покровов растения, где протекает фотосинтез. Таким образом, колленхима встречается только в надземных частях растительного организма. Ткань делится на три составляющие:

- Уголковая. Клетки имеют форму шестиугольников. Эта ткань утолщена неравномерно. Дело в том, что ее клеточки имеют более толстые стенки в углах. Уголковая колленхима характерна для двудольных растений.
- Пластинчатая. Встречается на молодых побегах многих видов деревьев. Ее клеточки напоминают параллелепипеды, вытянутые вдоль поверхности стебля.
- Рыхлая. На ранней стадии развития клетки этого типа покрова разъединяются в углах, после чего образуют межклетники.
Устройство склеренхимы
Этот вид покрова составляют омертвевшие клеточки. Склеренхиму можно встретить только у высших растений. В сравнении с колленхимой она способны выдерживать более высокие нагрузки. Стенки клеток склеренхимы пропитаны особым веществом — лигнином. Оно представляет собой смесь полимеров.
Склеренхима бывает двух типов:
Клеточная структура склереидов имеет одну важную особенность — стенки ее элементов одревеснели и часто дополнительно пропитаны кутином, кремнеземом либо известью. При этом склереиды делятся на 4 типа: каменистые, остеосклереиды, астросклереиды и палочковидные. К первому принадлежат клеточки, диаметр которых одинаков. Их можно встретить у плодов груши.

Остеосклереиды характеризуются расширенными концами клеток и встречаются, например, у чая. Палочкообразные клеточки характерны для бобовых. Астросклереиды приняли форму звезды и создают уникальный рисунок клеточной структуры. Эти клетки можно найти в листочках камелии.
Склеренхимные волокна имеют вытянутую форму и заострены на концах. Благодаря этому они могут располагаться на минимальном расстоянии друг от друга. Стенки их клеточек равномерно утолщены. Волокна встречаются в любом органе растительного организма. Они могут образовывать группы, составлять кольца либо равномерно распределены по проводящей ткани.
Уже из описания клеток, которые входят в состав механического покрова, можно точно сказать, какую функцию они выполняют. Все типы этой ткани растений предназначены для обеспечения целостности растительного организма. Благодаря особому строению клеток они обладают высокой эластичностью и прочностью.
Различные типы покровных тканей выполняют в некоторой мере и механические функции, защищая нежные ткани, расположенные ковнутри органа.
Но есть еще специальные клетки и слагаемые из них ткани, которые выполняют механические функции. Такие клетки и ткани называют механическими. Эти клетки и ткани усиливают противодействие всего растения или его органов, прежде всего излому или разрыву.
Механические клетки и ткани имеются во всех органах каждого растения, произрастающего в наземных условиях. В сильно растущих в длину частях растения, например в стеблях, элементы механических тканей также вытянуты, имеют прозенхимный характер. В органах со слабо выраженным ростом в длину механические клеточные элементы более или менее изодиаметричны — паренхимного характера. Такие элементы встречаются в плодах и листьях. Разумеется, абсолютно строгого разграничения между типами механических элементов не существует: нередко в стеблях имеются механические паренхимные клетки, а в листьях — вытянутые прозенхимные.
Все механические ткани по ряду признаков могут быть разделены на несколько групп, из которых каждая отличается определенными характерными чертами. Таких основных групп три: колленхима, лубяные и древесинные волокна и склереиды.
Колленхима встречается в стеблях, а также нередко в черешках и средних жилках листьев, реже в цветоножках и в плодоножках. Она находится в тех частях органов, где расположены сочные ткани, состоящие из живых клеток, и преимущественно свойственна двудольным растениям; у однодольных она встречается реже.
Колленхима служит для укрепления растущих органов. Она состоит из живых клеток, содержащих хлоропласты. Обычно колленхимные клетки длинные, иногда достигают значительной длины (2 мм), но среди них есть и вполне паренхимные клетки. Своеобразны утолщения оболочек колленхимных клеток. Наиболее часто наблюдается так называемое уголковое утолщение, когда полосы утолщений тянутся по углам клеток. Если утолщены тангентальные стенки клеток, то образуется пластинчатая колленхима. Характер утолщений колленхимы в значительной степени зависит от самого расположения клеток. Если клетки колленхимы расположены радиальными рядами, то утолщения возникают на тангентальных стенках. При беспорядочном расположении клеток утолщения концентрируются по углам.
Оболочки колленхимных клеток целлюлозные и на поперечных разрезах органов выделяются особым блеском. Особенность оболочек колленхимных клеток состоит в том, что в них содержится большое количество воды. Возможно, что это благоприятствует вытягиванию оболочки при росте органа.
Колленхима расположена почти всегда по периферии органа. Она дифференцируется раньше других механических тканей. Располагается колленхима или сплошным слоем в несколько рядов клеток вблизи периферии органа или собрана отдельными пучками, связанными с пучками проводящей системы. Если на стеблях есть ребра, то колленхима в виде компактных пучков тянется вдоль стебля по ребрам (например, зонтичные).
Колленхима принадлежит к числу простых тканей, т. е. клетки ее не смешиваются с клетками другого характера. Поэтому границы колленхимы обычно очерчены довольно резко. Однако иногда образуются переходные формы от колленхимных клеток к клеткам паренхимы.
Клетки колленхимы соприкасаются друг с другом тупо заканчивающимися концами и имеют простые поры. Клеточные полости их всегда широкие. Благодаря хлоропластам, колленхимные клетки, подобно соседним паренхимным клеткам коры, могут участвовать в фотосинтетической деятельности, но в значительно меньшей степени.
Лубяные волокна представляют собой вытянутые по оси органа длинные толстостенные клетки. Длина их у разных видов сильно колеблется. У конопли лубяные волокна имеют в среднем длину около 10 мм, у льна — около 40 мм, у крапивы — около 80 мм. Самые длинные волокна были найдены у субтропического прядильного растения рами — до 500 мм. У специальных текстильных сортов льна найдены лубяные волокна до 120 мм длины. У некоторых растений оболочки лубяных волокон остаются целлюлозными, но в большинстве случаев они более или менее рано одревесневают. Однако иногда одревеснение волокон происходит лишь у основания стебля.
К растениям с целлюлозными оболочками лубяных волокон принадлежат лен, рами. У конопли же, особенно у некоторых сортов ее, а также у крапивы оболочки лубяных волокон скоро одревесневают на всем протяжении стебля.
В молодом состоянии, во время роста, лубяные волокна содержат в своих полостях жизнедеятельный протопласт со многими ядрами. После окончательного формирования лубяного волокна протопласт обычно отмирает, остатки его лишь кое-где рассеяны в клеточной полости.
В живом протопласте лубяных волокон иногда обнаруживаются крахмальные зерна, но клеточная полость их обычно очень узка. Только у молодых волокон клеточная полость выражена отчетливо, у старых же, лишенных протопласта, полость почти совершенно исчезает. В толстых стенках иногда довольно хорошо заметна слоистость. Толщина оболочки даже у вполне сформировавшихся волокон не по всей длине одинакова: утолщенные участки перемежаются с утонченными, поэтому полость волокна местами суживается и расширяется.
У некоторых растений (виноградная лоза) полость лубяных волокон перегорожена тонкими перегородками. Такие анатомические элементы называются перегородчатыми лубяными волокнами.
Весьма характерно смыкание волокнистых клеток между собой.
Каждое волокно своими острыми концами вклинивается между другими волокнами. Такое взаимное соединение волокон, отличающееся от взаимного соединения паренхимных клеток, способствует наибольшей прочности ткани и происходит вследствие так называемого скользящего роста концов таких клеток.
На оболочках лубяных волокон имеются простые поры, расположенные по спирали. Последнее указывает на спиральную текстуру микрофибрилл во вторичной оболочке волокна. Спиральное расположение фибрилл придает волокну особую прочность. На спиральность текстуры указывает также то, что при механическом разрушении оболочки (сильное надавливание, приготовление срезов, поражение микроорганизмами) концы поврежденных оболочек иногда развертываются в виде вытянутой спирали.
Обычно лубяные волокна составляют простую ткань, т. е. к ним редко примешиваются другие анатомические элементы. Расположены лубяные волокна либо более или менее широким поясом, как в стеблях некоторых растений, либо собраны отдельными группами, образующими вместе с проводящими тканями сосудисто-волокнистые пучки.
Но есть и такие растения, у которых лубяные волокна не составляют отдельных пучков или компактных массивов, а перемешаны с паренхимными клетками. Такая паренхима называется лубяной.
При кажущейся простоте строения лубяные волокна по своей организации сильно варьируют даже у одного и того же растения. В нижней части стебля пучки лубяных волокон менее обособлены, чем в верхней части. Длина лубяных волокон в различных местах стебля одного и того же растения неодинакова: в нижней части расположены более короткие волокна, чем в средней и верхней частях стебля.
При произрастании растений одного и того же вида в различных климатических условиях создаются так называемые географические расы, своеобразные особенности которых отчасти передаются по наследству. У конопли, как и у других культурных растений, известно много географических рас. Расы конопли отличаются как количеством волокна (степенью волокнистости), так и качеством его. Анатомическое исследование обнаруживает значительную разницу и в строении лубяных волокон у географических рас конопли различного происхождения.
Лубяные волокна, подобные только что рассмотренным волокнам конопли, образуются перициклом — образовательной тканью, расположенной по периферии осевого органа, на границе с первичной корой. Лубяные волокна, возникшие в перицикле, называют первичными. Есть и вторичные лубяные волокна. Они возникают в результате деятельности другой образовательной ткани — камбия. В стеблях конопли, кроме первичных лубяных волокон, образуются и вторичные. То же наблюдается у кенафа, канатника, кендыря, рами — южных текстильных лубо-волокнистых растений. У льна и подсолнечника лубяные волокна только первичные.
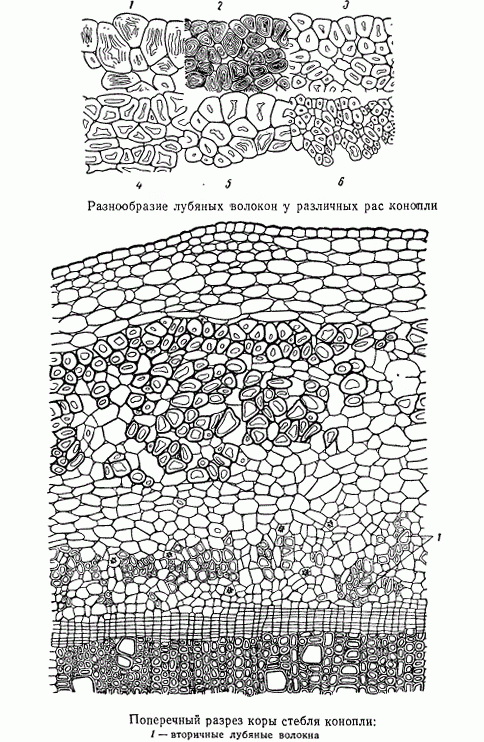
Вторичные лубяные волокна травянистых растений обычно развиты значительно слабее первичных. У конопли, например, вторичные лубяные волокна, по сравнению с первичными, имеют более мелкий просвет и одревесневшие оболочки, оболочки же первичных лубяных волокон многих сортов конопли остаются неодревесневшими или одревесневают весьма слабо. Текстильное значение имеют только первичные волокна и не только у конопли, но и у других, произрастающих в умеренном климатическом поясе травянистых текстильных растений. Вторичные лубяные волокна у этих растений наиболее обильно образуются лишь в базальной части стебля. Чем выше вдоль по стеблю, тем меньше развивается вторичных лубяных волокон; примерно на половине длины стебля они уже не образуются.
У древесных растений, наоборот, наиболее обильно развиты вторичные лубяные волокна. Первичные лубяные волокна, если и образуются, то лишь на ранней стадии развития, когда еще функционирует перицикл. Вторичные лубяные волокна в отличие от первичных, по-видимому, у всех древесных растений имеют одревесневшие оболочки. Примером вторичных лубяных волокон могут служить волокна липы, составляющие прочную основу так называемого лыка. Вторичные лубяные волокна никогда не достигают такой длины, как первичные.
Прочность лубяных волокон, особенно первичных, общеизвестна. Недаром из них выделывают различные ткани, канаты и пр. Волокна некоторых растений не уступают по прочности стали, далеко оставляя ее за собой по эластичности. Разумеется, колебания в прочности среди лубяных волокон тоже значительны и зависят от многих условий (степени одревеснения оболочек, влажности и пр.).
Больше всего лубяных волокон развивается в стеблях, в корнях их значительно меньше, часто не бывает совсем. У некоторых растений, преимущественно у однодольных (юкки, драцены, некоторые пальмы и пр.), хорошо развитые лубяные волокна встречаются в листьях. Есть лубяные волокна и в плодах различных типов (бобах, коробочках и др.).
Волокна, находящиеся в древесной части стебля, называются либриформом. Если лубяные волокна представляют собой самые толстостенные элементы коры, то ткань либриформа состоит из самых толстостенных анатомических элементов древесины. В основе своего строения либриформ сходен с лубяными волокнами и назначение его, по-видимому, такое же. Волокна либриформа значительно короче лубяных волокон (не больше 2 мм). Оболочки клеток либриформа всегда одревесневшие, снабжены простыми порами со щелевидными очертаниями. Так же, как и у лубяных волокон, имеющих простые поры, поры либриформа расположены по спирали. У некоторых растений (виноград) полость волокон либриформа, подобно полости лубяных волокон, перегорожена тонкими перегородками, не разделяющими, однако, окончательно самой клетки. Такие волокна либриформа, так же как и лубяные, называют перегородчатыми.
По своей структуре и внешнему виду клетки либриформа иногда представляют собой переход к другим типам тканей. Они могут быть очень короткими, приближаясь по форме к паренхиме. Оболочки таких клеток имеют простые поры, их протопласт долго остается живым, в них периодически накопляется крахмал. Но есть клетки либриформа, приближающиеся к водопроводящим элементам — трахеидам и сосудам. Оболочки таких клеток имеют окаймленные поры, протопласт в них скоро разрушается. Вообще тип либриформа не столь отчетлив, как тип лубяных волокон.
Либриформ также весьма распространен среди высших растений, как и лубяные волокна (твердый луб). У лиственных деревьев либриформ иногда занимает значительную часть древесины, в особенности в тех массивах ее, которые образовались во вторую половину вегетационного периода.
На примере либриформа особенно отчетливо выявляется общераспространенный принцип, обнаруживаемый при рассмотрении почти каждого анатомического элемента в растении, — принцип разнообразия функций, выполняемых той или другой клеткой или тканью. Либриформ наряду с основной функцией выполняет еще функции хранилища запасов и проведения воды. В соответствии с этим внешний вид и самая структура клеток либриформа не вполне устойчивы: в ряде случаев есть немало переходных форм, имеющих черты, характерные уже для других анатомических элементов.
Склереиды представляют собой клетки самой разнообразной формы, толстостенные, одревесневшие, не слишком большой длины или чаще паренхимные. Они встречаются в различных органах растений: плодах, листьях, стеблях. Наиболее типичным образцом склереид могут служить каменистые клетки, представляющие собой пример опорных элементов. В плодах груши и айвы каменистые клетки выполняют роль опоры для сочной мякоти зрелого плода. Каменистые клетки обычно паренхимные. Более вытянутые элементы встречаются в плодоножках. Твердые оболочки различных орехов, пергаментный слой в бобах также состоят из склереид. Нередко группы таких клеток способствуют раскрыванию плодов, разрыву более тонких стенок.
Иногда вытянутые склереиды ветвятся. Ветвистые склереиды чаще всего размещены одиночно и относительно велики. Неветвистые склереиды собраны группами, образуя подобие простой ткани. Нечто сходное происходит и с либриформом. Чаще всего клетки либриформа собраны группами, но нередко маленькие группы клеток либриформа или даже отдельные клетки перемежаются с клетками другого характера.
Такие ткани, состоящие из разнообразных анатомических элементов, можно назвать смешанными.
Механические ткани в каждом органе сконцентрированы таким образом, что обусловливают прочность на разрыв или изгиб в определенном направлении. В стебле, подвергающемся преимущественно изгибу, механические ткани расположены по периферии, что особенно ясно выражено у травянистых растений.
В корнях, подвергающихся преимущественно растяжению, наоборот, периферическая зона (кора) почти лишена механических элементов, и они расположены ближе к центру оси корня.
На основе анализа закономерностей в распределении механических тканей в растениях некоторые исследователи разработали ряд схем конструкций органов растений с использованием строительно-механических принципов. Следует, однако, признать, что строительно-механический принцип объясняет лишь одну сторону функционирования системы механических тканей.
Дело в том, что механические анатомические элементы редко выполняют только механические функции. Только с этих позиций трудно объяснить, например, значение незначительного по толщине кольца лубяных волокон в древесном стволе с его солидной древесиной и притом нередко весьма прочной.
Структура и функции каждого органа сложны и разнообразны, в силу чего с помощью чисто механических схем нельзя выяснить закономерности в организации структуры растений, тем более что между различными анатомическими элементами существует ряд промежуточных форм. Структура пластична. В живом организме структура — процесс.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
Читайте также:
- Какие права у ребенка инвалида
- В каком случае закон независимого наследования признаков несправедлив
- Что такое право и как оно влияет на общественные отношения соедини пары
- К чему снится воровство у покойника
- Как обеспечить безопасность дошкольников конспекты занятий по основам безопасности детей дошкольного возраста