
Как называются признаки детерминированные несколькими неаллельными генами и их тип наследования
Обновлено: 30.06.2024
Основные понятия генетики
Генетический анализ
Гибридологический метод
Взаимодействие генов
Тестовые задания
Основные понятия генетики
Наследственность — свойство организмов сохранять генетическую информацию, детерминирующую признаки, особенности развития и жизнедеятельность потомков. Наследственность закрепляет признаки, характерные для вида.
Наследование — способ передачи наследственной информации от одного поколения к другому.
Изменчивость — свойство организмов изменять признаки, полу-ченные от родителей, или приобретать новые в процессе индивиду-ального развития.
Ген — единица наследственности. В структурном отношении — это определенная последовательность нуклеотидов ДНК, кодирующая структуру полипептидной цепи. В функциональном отношении (как единица функции — цистрон) — это совокупность кодирующих и регуляторных последовательностей, одновременно участвующих в процессе транскрипции.
Гены располагаются в хромосомах линейно. Участок хромосомы, в котором расположен определенный ген, называется локусом. Гены одной хромосомы образуют группу сцепления и наследуются вместе. При кроссинговере сцепление генов нарушается.
Аллель — одно из возможных состояний гена (конкретное состо-яние кодирующей последовательности: определенный состав, по-следовательность и количество нуклеотидов в локусе).
Аллельные гены — гены, определяющие развитие одного и того же признака (цвет) в одинаковых или разных фенотипических про-явлениях (желтый, зеленый); локализованы в одинаковых локусах гомологичных хромосом.
Неаллельные гены — гены, определяющие развитие разных при-знаков (цвет и форма). Могут быть локализованы в гомологичных хромосомах, но в разных локусах, или в негомологичных хромосомах.
Гомозигота — организм, в гомологичных хромосомах которого находятся одинаковые аллели одного гена (изоаллели) — аа, АА, ААВВ, ааВВ; образует один тип гамет.
Гетерозигота — организм, в гомологичных хромосомах которого локализованы разные аллели одного гена (Аа, АаВв); образует разные типы гамет.
Гемизигота — организм, содержащий только один аллель определенного гена (мужской организм гемизиготен по генам, локализованным в X- и Y-хромосомах). Гены, расположенные в участке Y-хромосомы, не гомологичном Х-хромосоме, называются сцепленными с Y-хромосомой, а наследование — голандрическим.
Гибридизация — скрещивание гомозиготных особей, отличающихся по генотипу (фенотипу).
Рецессивный аллель — аллель, проявляющийся только у гомозиготных организмов (аа); у гетерозиготных подавляется доминантным аллелем.
Доминантный аллель — аллель, проявляющийся у гомо- и у ге-терозиготных организмов (АА, Аа).
Фенотип — совокупность всех признаков, свойств, особенностей развития и жизнедеятельности организма. Это результат реализации генотипа в конкретных условиях среды.
Генотип — система всех взаимодействующих аллелей организма.
Геном — количество генов в гаплоидном наборе, характерное для вида.
Генетический анализ
Генетический анализ — совокупность методов исследования на-следственных свойств организма, определяемых его генотипом. Анализ элементов генотипа (групп сцепления, генов и внутриген- ных структур) осуществляется, как правило, по фенотипу (опосредованно по признакам). Генетический анализ является по существу анализом признаков, контролируемых генотипом. Признаки могут наследоваться моногенно (один ген — один признак, один ген — несколько признаков) и полигенно (несколько генов — один признак). При всех типах наследования среда может оказывать определенное влияние на проявление признака.
В зависимости от задач и особенностей изучаемого объекта генетический анализ проводят на популяционном, организменном, клеточном и молекулярном уровнях. К основным методам генетического анализа относятся: гибридологический, близнецовый, биохимический, цитогенетический, молекулярно-генетический, популяционно-статистический и др.
Наиболее удобные биологические объекты для генетических ис-следований (муха дрозофила, горох, дрожжи, бактерии и др.) имеют целый ряд особенностей, которые позволяют проводить гибридизацию и последующий анализ потомства: скрещивание в искусственных условиях, достаточно высокая плодовитость, использование ста-тистических методов при анализе потомства, быстрая смена поколений, небольшое число групп сцепления в геноме, незначительные изменения признаков под влиянием окружающей среды.
Моногибридное скрещивание
Моногибридным называется скрещивание, при котором просле-живается наследование одного признака.
Первый закон Менделя — закон единообразия гибридов первого поколения: при скрещивании гомозиготных особей, отличающихся альтернативным проявлением одного признака, все потомство будет единообразным по фенотипу и генотипу. Полученные особи называются гибридами.
Второй закон Менделя — закон расщепления: при скрещивании гибридов первого поколения между собой в потомстве происходит рас-щепление признака по фенотипу 3 :1, а по генотипу 1:2:1.
Эти соотношения при расщеплении можно получить при следу-ющих условиях:
— число потомков должно быть большим, так как это статистиче-ская закономерность;
— между аллелями имеется полное доминирование.
При неполном доминировании расщепление по генотипу и фенотипу совпадает 1:2:1.
Изучая моногибридное скрещивание, Г. Мендель разработал разные типы скрещивания, в том числе и анализирующее.
Анализирующее скрещивание используют для выяснения генотипа исследуемого организма: организм с доминантным признаком скрещивают с рецессивной гомозиготой и анализируют потомство. При этом возможны два варианта:
— все потомство единообразно — организм с доминантным при-знаком гомозиготен;
— потомки различаются фенотипически — организм с доми-нантным признаком гетерозиготен.
Дигибридиое скрещивание
Дигибридное скрещивание — это скрещивание, при котором ана-лизируется наследование двух признаков одновременно. Мендель показал, что закон единообразия гибридов первого поколения справедлив для любого количества признаков, в том числе и для дигибридного скрещивания.
Третий закон Менделя — закон независимого наследования и независимого комбинирования признаков: при ди- и полиги- бридных скрещиваниях каждый признак наследуется независимо от другого, расщепляясь в соотношении 3:1. При дигибридном скрещивании в F2 формируются четыре фенотипа в соотношении 9:3 :3 :1, при этом два из них — рекомбинантные. Закон соблюдается, если неаллельные гены находятся в разных парах гомологичных хромосом и отсутствует взаимодействие между ними.
Цитологические основы законов Менделя базируются на процессах гаметогенеза и оплодотворения.
Законы Менделя выполняются при определенных условиях.
Условия менделирования признаков:
• моногенное наследование;
• полное доминирование;
• равновероятное образование всех гамет;
• равновероятная встреча всех гамет при оплодотворении;
• равновероятная выживаемость всех зигот;
• отсутствие летальных мутаций;
• отсутствие взаимодействий между неаллельными генами (при полигибридном скрещивании);
• полная пенетрантность гена;
• выраженная стойкая экспрессивность гена.
Взаимодействие генов
Генотип — это система взаимодействующих аллелей. Эти взаимо-отношения могут быть представлены в виде взаимодействия аллельных и неаллельных генов. Один ген может бьггь представлен двумя (чаще) или большим числом аллелей (множественные аллели), но в диплоидном организме содержится только два аллеля одного гена.
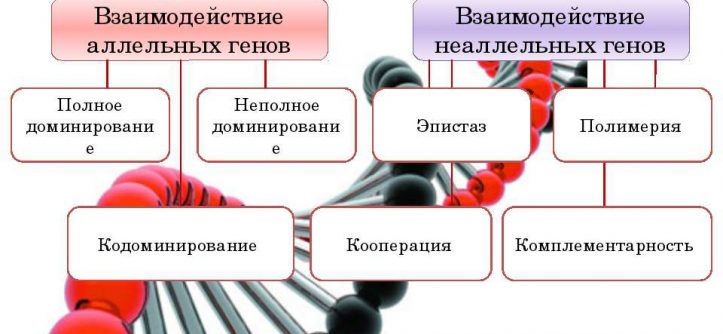
Взаимодействие аллельных генов
Взаимодействие аллельных генов — это взаимодействие между аллелями одного и того же гена. Взаимодействие осуществляется по типу: полного доминирования, неполного доминирования, сверхдо-минирования, кодоминирования.
Полное доминирование — доминантный аллель полностью подавляет рецессивный; проявление доминантного аллеля не зависит от присутствия в генотипе другого аллеля; гетерозиготы и гомозиготы фенотипически не отличаются (желтый цвет горошин доминирует над зеленым, карие глаза — над голубыми).
Неполное доминирование — доминантный аллель неполностью подавляет рецессивный, а у гетерозигот формируется промежуточный фенотип (розовые цветки ночной красавицы).
Кодоминирование — два доминантных аллеля одного гена про-являются в фенотипе независимо друг от друга (организм с генотипом IA IB определяет синтез двух видов антигенов А и В).
Сверхдоминирование — у гетерозигот признак выражен сильнее, чем у гомозигот; доминантный аллель в гетерозиготном состоянии имеет более сильное проявление, чем в гомозиготном (гетерозисная сила).
Взаимодействие неаллельных генов
При моногенном наследовании один ген определяет развитие одного признака, но иногда для формирования одного признака необходимо несколько генов, такой тип наследования называется по- лигенным. Неаллельные гены взаимодействуют друг с другом, при этом у потомков наблюдаются новые фенотипы в и необычное расщепление в F2, которое представляет собой видоизменение классической формулы наследования, так как признак не менделирует. Различают три основных типа взаимодействия неаллельных генов: комплементарность, эпистаз (качественные признаки), полимерия (количественные признаки).
Комплементарность — тип взаимодействия, при котором два не-аллельных гена, обычно доминантных, находясь одновременно в генотипе, приводят к формированию нового проявления признака. У человека для образования некоторых признаков необходимо ком-плементарное взаимодополняющее действие генов (формирование нормального слуха, синтез гемоглобина).
Эпистаз — тип взаимодействия, при котором наблюдается пода-вление действия одного гена другим, неаллельным ему. Подавляющий ген называется эпистатическим геном (геном супрессором или ингибитором), а подавляемый — гипостатическим. Различают эпистаз доминантный (доминантный аллель одного гена подавляет проявление доминантных аллелей другого гена) и рецессивный (рецессивный аллель подавляет проявление доминантных аллелей другого гена только в гомозиготном состоянии).
Полимерия — тип взаимодействия неаллельных генов, когда при формировании признака важно количество доминантных генов, влияющих на степень его выраженности. Гены обозначаются одинаковыми буквами (цифровое обозначение с нижним индексом), чтобы подчер-кнуть однонаправленность их действия А], А2, . А10. У человека по типу полимерии наследуются пигментация кожи (чем больше доминантных аллелей, тем больше образуется меланина и тем интенсивнее окраска кожи), а также большинство количественных признаков, кото-рые можно измерить или подсчитать (вес, рост, артериальное давление).
С точки зрения генетики, наиболее изученной является система АВ0, определяющая I (0), II (А), III (В) и IV (АВ) группы крови. На поверхности эритроцитов могут находиться агглютиногены (антигены) А и В, а в плазме крови – агглютинины (антитела) и . В норме одноименные агглютиногены и агглютинины совместно не обнаруживаются. Нужно отметить, что А- и В-антигены образуют многочисленный ряд антигенов (А1, А2 . A; В1, В2 …В).
Понятие о комплиментарном взаимодействии генов. Примеры.
Эпистатическое взаимодействие генов. Примеры.
Подавляющий ген – эпистатический
Подавляемый ген – гипостатический
Угнетение могут вызывать как доминантные, так и рецессивные гены (А> В, а> В, В> А, В> А), и в зависимости от этого различают эпистаз доминантный и рецессивный. Подавляющий ген получил название ингибитора или супрессора. Гены-ингибиторы в основном не детерминируют развитие определенного признака, а лишь подавляют действие другого гена.
Ген, эффект которого подавляется, получил название гипостатичного. При эпистатичном взаимодействии генов расщепление по фенотипу в F2 составляет 13:3; 12:3:1 или 9:3:4 и др. Окрас плодов тыквы, масть лошадей определяются этим типом взаимодействия.
Если ген-супрессор рецессивный, то возникает криптомерия (греч. хриштад - тайный, скрытый).
У человека таким примером может быть "Бомбейский феномен". В этом случае редкий рецессивный аллель "h" в гомозиготном состоянии (hh) подавляет активность гена jB (определяющий В (III) группу крови системы АВО). Поэтому женщина с генотипом jв_hh, фенотипно имеет I группу крови - 0 (I).
При эпистазе один из генов (В) выражается фенотипически лишь при отсутствии в генотипе определенного аллеля другого гена (А). В его присутствии действие гена В не проявляется. В строгом смысле слова, этот вид взаимодействия неаллельных генов может быть рассмотрен как вариант взаимодополняющего действия определенных аллелей этих генов, когда один из них способен обеспечить развитие признака, но лишь в присутствии определенного аллеля другого гена. В этой ситуации фенотип организма зависит от конкретного сочетания аллелей неаллельных генов в их генотипах и расщепление по фенотипу в потомстве двух дигетерозигот по этим генам может быть различным.
При доминантном эпистазе, когда доминантный аллель одного гена (А) препятствует проявлению аллелей другого гена (В или b), расщепление в потомстве зависит от их фенотипического значения и может выражаться соотношениями 12:3:1 или 13:3 (рис. 6.19). При рецессивном эпистазе ген, определяющий какой-то признак (В), не проявляется у гомозигот по рецессивному аллелю другого гена (аа). Расщепление в потомстве двух дигетерозигот по таким генам будет соответствовать соотношению 9:3:4 (рис. 6.20). Невозможность формирования признака при рецессивном эпистазе расценивают также как проявление несостоявшегося комплементарного взаимодействия, которое возникает между доминантным аллелем эпистатического гена и аллелями гена, определяющего этот признак.
Полимерия и ее роль в детерминации количественных признаков. Аддитивный эффект.
Важная особенность полимерии - суммация действия неаллельних генов на развитие количественных признаков. Если при моногенном наследовании признака возможны три варианта "доз" гена в генотипе: АА, Аа, аа, то при полигенных количество их возрастает до четырех и более. Суммация "доз" полимерных генов обесчивает существования непрерывных рядов количественных изменений.
Биологическое значение полимерии заключается еще и в том, что признаки, кодируемые этими генами, более стабильны, чем те, которые кодируются одним геном. Организм без полимерных генов был бы очень неустойчивым: любая мутация или рекомбинация приводила бы к резкой изменчивости, а это в большинстве случаев имеет неблагоприятный характер.
У животных и растений есть много полигенных признаков, среди них и ценные для хозяйства: интенсивность роста, скороспелость, яйценоскость, количество молока, содержание сахаристых веществ и витаминов и т.п.
Пигментация кожи у человека определяется пятью или шестью полимерными генами. В коренных жителей Африки (негроидной расы) преобладают доминантные аллели, у представителей европеоидной расы - рецессивные. Поэтому мулаты имеют промежуточную пигментацию, но при браках мулатов у них возможно появление как более, так и менее интенсивно пигментированных детей.
Многие морфологические, физиологические и патологические особенности человека определяются полимерными генами: рост, масса тела, величина артериального давления и др. Развитие таких признаков у человека подчиняется общим законам полигенного наследования и зависит от условий среды. В этих в случаях наблюдается, например, склонность к гипертонической болезни, ожирению и др. Данные признаки при благоприятных условиях среды могут не проявиться или проявиться незначительно. Эти полигенные признаки отличаются от моногенных. Изменяя условия среды можно обеспечить профилактику ряда полигенных заболеваний.
Наследование признаков при полимерном взаимодействии генов. В том случае, когда сложный признак определяется несколькими парами генов в генотипе и их взаимодействие сводится к накоплению эффекта действия определенных аллелей этих генов, в потомстве гетерозигот наблюдается разная степень выраженности признака, зависящая от суммарной дозы соответствующих аллелей. Например, степень пигментации кожи у человека, определяемая четырьмя парами генов, колеблется от максимально выраженной у гомозигот по доминантным аллелям во всех четырех парах (Р1Р1Р2Р2Р3Р3Р4Р4) до минимальной у гомозигот по рецессивным аллелям (р1р1р2р2р3р3р4р4). При браке двух мулатов, гетерозиготных по всем четырем парам, которые образуют по 2 4 = 16 типов гамет, получается потомство, 1/256 которого имеет максимальную пигментацию кожи, 1/256 — минимальную, а остальные характеризуются промежуточными показателями экспрессивности этого признака. В разобранном примере доминантные аллели полигенов определяют синтез пигмента, а рецессивные — практически не обеспечивают этого признака. В клетках кожи организмов, гомозиготных по рецессивным аллелям всех генов, содержится минимальное количество гранул пигмента.
Плейотропное действие генов. Примеры.
Назовите основные биометрические характеристики, испльзуемые при генетико-математическом анализе количественных признаков.
Физиологические — относятся к форме тела. В качестве примера можно привести: отпечатки пальцев, распознавание лица, ДНК, ладонь руки, сетчатка глаза, запах, голос.
Поведенческие — связаны с поведением человека. Например, походка и речь. Порой используется термин англ. behaviometrics для этого класса биометрии.
Понятие о варианте и вариационном ряде.
Основные обозначения вариационного ряда
р — частота ("вес") варианты, число ее повторений в вариационном ряду;
n — общее число наблюдений (т.е. сумма всех частот, n = Σр);
Vmax и Vmin — крайние варианты, ограничивающие вариационный ряд (лимиты ряда);
А — амплитуда ряда (т.е. разность между максимальной и минимальной вариантами,А = Vmax — Vmin)
Виды вариаций:
а) простой — это ряд, в котором каждая вариата встречается по одному разу (р=1);
6) взвешенный — ряд, в котором отдельные варианты встречаются неоднократно (с разной частотой).
Сущность средней арифметической, среднего квадратического отклонения, дисперсии и методы их расчета.
для оценки состояния здоровья — например, параметров физического развития (средний рост, средняя масса тела, среднее значение жизненной емкости легких и др.), соматических показателей (средний уровень сахара в крови, средняя величина пульса, средняя СОЭ и др.);
для оценки организации работы лечебно-профилактических и санитарно-противоэпидемических учреждений, а также деятельности отдельных врачей и других медицинских работников (средняя длительность пребывания больного на койке, среднее число посещений на 1 ч приема в поликлинике и др.);
для оценки состояния окружающей среды.
Суммировать варианты: V1+V2+V3+. +Vn = Σ V;
Сумму вариант разделить на общее число наблюдений: М = Σ V / n
Получить произведение каждой варианты на ее частоту — Vp
Найти сумму произведений вариант на частоты: V1p1 + V2p2+ V3p3 +. + Vnpn = Σ Vp
Полученную сумму разделить на общее число наблюдений: М = Σ Vp / n
Среднее квадратичное отклонение определяется как обобщающая характеристика размеров вариации признака в совокупности. Оно равно квадратному корню из среднего квадрата отклонений отдельных значений признака от средней арифметической, т.е. корень из дисперсии и может быть найдена так:
1. Для первичного ряда:

2. Для вариационного ряда:

Преобразование формулы среднего квадратичного отклонени приводит ее к виду, более удобному для практических расчетов:

Среднее квадратичное отклонение определяет на сколько в среднем отклоняются конкретные варианты от их среднего значения, и к тому же является абсолютной мерой колеблемости признака и выражается в тех же единицах, что и варианты, и поэтому хорошо интерпретируется.
Найти отклонение (разность) каждой варианты от среднеарифметической величины ряда (d = V — М);
Возвести каждое из этих отклонений в квадрат (d2);
Получить произведение квадрата каждого отклонения на частоту (d2р);
Найти сумму этих отклонений: d21p1 + d22p2 + d23p3 +. + d2npn = Σ d2р;
Полученную сумму разделить на общее число наблюдений (при n Фенотипическая дисперсия - статистическая величина, с помощью которой оценивается размах вариативности какого-либо количественного признака в популяции.
В привычном представлении второй закон Менделя определяет возможные комбинации из двух аллелей одного гена и дает оценку частот таких комбинаций. Напомним, в случае скрещивания двух носителей рецессивной мутации A/a ожидается классическое расщепление генотипов в потомстве:
25% A/A 50% A/a 25% a/a
Такой случай называется моногибридным скрещиванием, поскольку в нем отслеживается судьба одного гена.
Мендель не знал про хромосомы (их открыли через 20 лет после того, как он опубликовал свои законы), но мы-то знаем, что наблюдаемое расщепление возникает оттого, что от каждого из родителей потомству передается одна хромосома из каждой пары. Так что фактически закон Менделя описывает, как наследуются хромосомы от каждого из родителей.
Независимое наследование генов
Каждый организм несет тысячи генов, которые могут содержать мутации и наследуются все одновременно. Понятно, что чем больше признаков, определяемых разными генами, мы хотим отследить, тем больше комбинаций они формируют.
Рассмотрим пример наследования двух генов. Допустим, генотип обоих родителей A/a; B/b. В этом случае A и B - два разных, независимо наследуемых гена. Видно, что добавление всего одного гена существенно усложняет анализ, как показано на рисунке.
Число возможных генотипов у потомства в таком случае равно числу цветов в приведенной таблице.
Например, генотип A/a; B/b при независимом наследовании ожидается в четырех случаях из 16 (25%). Генотип a/a; b/b будет появляться только в 1 случае из 16 (6,25%).
С биологической точки зрения независимое наследование означает, что гены A и B находятся в разных парах хромосом. Приведем конкретный пример.

Гены A и B находятся в разных хромосомных парах и поэтому их аллели передаются потомству независимо друг от друга. Поэтому в приведенном примере всего лишь одна пара независимо наследующихся генов дает целых 9 возможных вариантов потомков!
У собак ген MKLN1, мутация в котором является причиной летального акродерматита у бультерьеров, находится в 14-й паре хромосом. Ген PKD1, мутация в котором приводит к поликистозу почек в этой породе, расположен в 6-й паре хромосом. Эти гены наследуются независимо друг от друга.
Напомним, что у собаки 39 пар хромосом, у кошки - 19 пар, у человека - 23. Таким образом, у собак мы теоретически можем одновременно отслеживать не более 39 независимо наследуемых генов, по числу пар хромосом. Таблица распределения генотипов в потомстве получится в этом случае уже совсем сложной!
*********************
Заметьте, что при записи генотипа мы разделили гены A и B знаком точка с запятой. В классической генетике так обозначают гены, расположенные в разных парах хромосом.
*********************
Сцепленное наследование генов
Но как же наследуются гены, расположенные в одной и той же паре хромосом? Поскольку каждая хромосома наследуется целиком, эти гены будут также наследоваться вместе, как показано на рисунке.
В приведенном примере и отец и мать - гетерозиготы по генам C и D, то есть у каждого из них есть и доминантные аллели (C и D) и рецессивные аллели (c и d). Однако в этом примере мы рассмотрели случай, когда у отца оба доминантных аллеля находятся в одной хромосоме из пары (C,D), а у матери - в разных (C,d и c,D), как показано на Рисунке 2.
Понятно, что у отца и у матери аллели могут быть распределены по хромосомам и одинаковым образом, но в этом случае и генотипы потомства будут иными.
Поскольку гены C и D расположены в одной хромосоме, то есть в одной непрерывной молекуле ДНК, они не могут наследоваться независимо, как в случае рассмотренного выше случае для генов A и B!

Гены C и D находятся в одной хромосоме и поэтому их аллели передаются потомству вместе, ведь они физически находятся в одной молекуле ДНК.
В половине сперматозоидов отца будет содержаться два доминантных аллеля C и D, а в половине - два рецессивных. У матери возможно формирование также двух типов яйцеклеток - C,d и c,D. Гены C и D в генетике называют сцепленными, поскольку их наследование нарушает расщепление генотипов потомства, наблюдаемое для независимо наследуемых генов.
******************
Обратите внимание, что аллели генов, расположенные в одной хромосоме из пары, записываются через запятую (например: C,d). Хромосомы в паре разделяются косой чертой (например: C,d/c,d)
******************
Нарушается ли второй закон Менделя в случае сцепленных генов? Конечно, нет! И в том и в другом случае речь идет о наследовании хромосом: одна хромосома в каждой паре достается от отца, а вторая - от матери. Понятно, что отличия хромосом в каждой паре могут затрагивать не один ген, но их передача потомству происходит по тем же правилам. В случае сцепленных генов отличие от моногибридного скрещивания состоит в том, что с каждой хромосомой передаются варианты многих генов - в приведенном примере это аллели генов C и D.
♦ Свойства генов и особенности их проявления в признаках:
■ ген дискретен в своем действии, т.е. обособлен в своей активности от других генов;
■ один ген отвечает за проявление одного строго определенного признака или нескольких признаков (плейотропия);
■ один признак может быть результатом действия нескольких генов (аллельных или неаллельных);
■ ген может усиливать степень проявления признака при увеличении числа его доминантных аллелей;
■ ген может взаимодействовать с другими генами; это приводит к появлению новых признаков;
■ изменение положения гена в хромосоме или влияние факторов внешней среды могут модифицировать его проявление в признаках;
■ ген обладает способностью к мутациям.
Взаимодействие аллельных генов
Взаимодействие генов — явление, когда за один признак отвечает несколько генов (или аллелей).
Аллельное взаимодействие — это взаимодействие аллелей одного и того же гена (за признак отвечает несколько аллелей одного и того же гена).
❖ Типы аллельных взаимодействий:
■ доминирование,
■ неполное доминирование,
■ сверхдоминирование,
■ кодоминирование.
Доминирование — тип взаимодействия двух аллелей одного гена, когда один (доминантный) из них полностью исключает действие другого (рецессивного). Примеры: доминирование у человека темных волос над светлыми, карих глаз над голубыми.
Неполное доминирование — степень активности доминантного аллеля недостаточна для того, чтобы полностью подавить действие рецессивного аллеля и обеспечить полное проявление доминантного признака.
■ В этом случае у гетерозигот формируется промежуточный (по отношению к родительским признакам) признак — имеет место промежуточный характер наследования. Этот признак будет наблюдаться у гибридов первого поколения и гетерозигот второго поколения. Во втором поколении расщепление по фенотипу и генотипу оказывается одинаковым 1:2:1 (одну часть составляет доминантная гомозигота АА с выраженным доминантным признаком, две части составляет гетерозигота Аа с промежуточным признаком и одну часть составляет гомозигота аа с рецессивным признаком).

■ Примеры неполного доминирования: наследование формы (курчавости) волос у человека, масти крупного рогатого скота, окраски цветков у растения ночная красавица (см. таблицу).
Сверхдоминирование — более сильное проявление признака у гетерозиготной особи (Аа), чем у любой из гомозигот (АА и аа).
Кодоминирование — оба аллеля равноценны, не подавляют друг друга и участвуют в определении признака у гетерозиготной особи. Пример: наследование IV группы крови у человека, которая детерминируется одновременным присутствием в генотипе двух кодоминантных генов I А и I в . Первый из этих генов детерминирует синтез в эритроцитах белка-антигена А, второй — синтез белка-антигена В; наличие обоих этих генов в генотипе приводит к тому, что у людей с IV группой крови эритроциты содержат как белок-антиген А, так и белок-антиген В.
Взаимодействие неаллельных генов
Неаллельное (или межаллельное) взаимодействие — это взаимодействие аллелей разных генов, т.е. генов, располагающихся в негомологичных хромосомах или разных локусах гомологичных хромосом.
■ Неаллельное взаимодействие генов приводит к модификации менделевского расщепления по фенотипу 9 : 3 : 1, т.е. к появлению в потомстве гетерозиготы иных расщеплений, например 9 : 3 : 4; 9 : 6 : 1; 12 : 3 : 1 и др.
❖ Основные типы межаллельных взаимодействий:
■ комплементарность;
■ эпистаз;
■ полимерия.
Замечание: комплементарное и эпистатическое взаимодействия возникают в тех случаях, когда признак контролируется одной парой неаллельных генов.
Комплементарное, или дополнительное, взаимодействие — такой тип межаллельного взаимодействия генов, при котором одновременное присутствие в генотипе гибрида доминантных генов разных аллельных пар приводит к появлению нового признака, отсутствующего у обоих родителей.
Пример: наследование окраски цветков душистого горошка (родительские растения с генотипами А-вв, ааВ- имеют белые цветки, гибриды с генотипом А-В- пурпурные; см. таблицу).

Объяснение: пурпурный пигмент образуется с помощью специального фермента, который синтезируется только при наличии обоих доминантных генов: как А, так и В. Цветки родительских особей имеют белый цвет, так как в генотипе каждого из них присутствует только один из этих генов.
Во втором поколении при самоопылении, обеспечивающем равновероятное (случайное) образование гамет и зигот разного типа, наблюдается расщепление по фенотипу в соотношении пурпурной и белой окраски цветков как 9 : 7 (9 пурпурных: A-В- и 7 белых: ЗА-Bb, ЗааВ-. 1aabb).
Эпистаз — такой тип межаллельного взаимодействия генов, при котором аллели одного гена подавляют проявление аллельной пары другого гена, и подавляемый признак не проявляется.
Супрессор (или ген-ингибитор) — ген, подавляющий действие других неаллельных генов. Супрессором может быть как доминантный, так и рецессивный ген.
Доминантный эпистаз — эпистаз, в котором супрессором является доминантный ген. При доминантном эпистазе во втором поколении наблюдается расщепление фенотипа 12:3:1 или 13:3.
Рецессивный эпистаз (криптомерия) — эпистаз, в котором супрессором является рецессивный ген. При рецессивном эпистазе во втором поколении наблюдается расщепление по фенотипу 9:3:4.
Пример эпистаза: наследование окраски шерсти у домашних кроликов. Синтез черного пигмента детерминирует рецессивный ген с, доминантная аллель I другого гена является супрессором, подавляя действие гена с. Тогда кролики с генотипами C-I-, ccl-будут белыми, кролики с генотипами C-ii — серыми, а с генотипом ссii — черными.
Многие признаки контролируются двумя и более парами неаллельных генов (называемых в этом случае полимерными).
Полимерия — взаимодействие нескольких неаллельных полимерных генов. При полимерии степень выраженности фенотипического признака часто зависит от числа полимерных генов, ответственных за его проявление. При кумулятивной полимерии действие генов суммируется; примеры: масса тела, молочность крупного рогатого скота, яйценоскость кур, некоторые параметры умственных способностей человека и др. При некумулятивной полимерии степень проявления признака от числа доминантных генов в генотипе не зависит (пример: оперенность ног у кур).
Плейотропия — зависимость нескольких признаков от одного гена. Каждый плейотропный ген оказывает какое-то основное действие, но модифицирует проявление других генов.
Сцепление генов. Опыты Моргана
Сцепленные гены — любые гены, расположенные в одной хромосоме.
Группа сцепления — все гены, расположенные в одной хромосоме.
■ Количество групп сцепления равно числу пар хромосом (т.е. гаплоидному числу хромосом). У человека 46 хромосом, т.е. 23 группы сцепления.
■ Наследование признаков, за которые отвечают гены из одной группы сцепления, не подчиняется законам Менделя.
Опыты Т. Моргана (1911-1912 гг.): анализ наследования двух пар альтернативных признаков у мух-дрозофил — серого (В) и черного (b) цвета тела и нормальной (V) или укороченной (v) длины крыльев.
Первая серия опытов: скрещивание гомозиготной доминантной (BBVV) особи (с серым цветом тела и нормальной длиной крыльев) с гомозиготной рецессивной (bbvv) особью черного цвета с короткими крыльями. Все потомки F1, в соответствии с первым законом Менделя, являются доминантными гетерозиготными (BbVv) особями серого цвета с нормальными крыльями.
Вторая серия опытов: анализирующее скрещивание гибридов первого поколения — гомозиготной рецессивной (черной короткокрылой) самки (bbvv) с дигетерозиготным (серым с нормальными крыльями) самцом (BbVv). Если предположить, что два гена, относящиеся к разным аллельным парам, локализованы в разных хромосомах, то у дигетерозиготы следует ожидать образования (в равных количествах) четырех типов гамет: BV, bV, Bv и bv. Тогда, согласно третьему закону Менделя, в потомстве должны присутствовать четыре разных фенотипа в равном количестве (по 25%). В действительности присутствовали только два фенотипа (в соотношении 1 : 1).
■ Это означает, что доминантные гены В и V, относящиеся к разным аллельным парам, локализованы в одной хромосоме (из пары гомологичных хромосом) и попадают в одну гамету, а оба рецессивных гена в и v локализованы в другой хромосоме и вместе попадают в другую гамету. Поэтому у дигетерозиготного самца мухи-дрозофилы образуется не четыре типа гамет (когда гены расположены в разных хромосомах), а только два: BV (50%) и bv (50%), и, следовательно, потомки F2 будут иметь два сочетания признаков.
Третья серия опытов: проверка предположения о полном сцеплении генов путем анализирующего скрещивания дигетерозиготной (серой с нормальными крыльями) самки (BbVv) из поколения F1 с гомозиготным рецессивным (черным короткокрылым) самцом (bbvv) из родительского поколения. В результате были получены потомки четырех фенотипов в следующем соотношении: по 41,5% особей с серым телом и нормальными крыльями (генотип BbVv) и особей с черным телом и короткими крыльями (генотип bbvv), и по 8,5% серых короткокрылых особей (генотип Bbvv) и черных особей с нормальными крыльями (генотип bbVv).
Отсюда следует, что сцепленные гены, т.е. гены, локализованные в одной хромосоме, не всегда передаются вместе, т.е. сцепление может быть неполным. Это связано с явлением кроссинговера. вероятность которого в данном случае составляет 17%.
Кроссинговер
Кроссинговер — явление обмена участками гомологичных хрома-тид в процессе их конъюгации в профазе мейоза I.
■ У гетерозиготных организмов кроссинговер приводит к пере-комбинации генетического материала.
■ Кроссинговер происходит не всегда; его частота зависит от расстояния между генами (подробнее о расстоянии между генами см. ниже).
■ Кроссинговер имеет место у всех растений и животных, за исключением самца мухи-дрозофилы и самки тутового шелкопряда.
■ Значение кроссинговера: он позволяет создавать новые комбинации генов и тем самым повысить наследственную изменчивость, необходимую для расширения возможностей адаптации организмов к изменившимся условиям среды.
Рекомбинация — появление новых сочетаний генов в результате кроссинговера, свободного комбинирования хромосом при образовании гамет или при их слиянии в ходе оплодотворения.
Кроссоверные (или рекомбинантные) особи — особи, образующиеся из гамет с новым, полученным путем кроссинговера, сочетанием аллелей.
Закон Моргана (закон сцепленного наследования): гены, расположенные в одной хромосоме (т.е. входящие в одну группу сцепления), наследуются преимущественно, т.е. с наибольшей вероятностью, вместе (сцепленно).
Хромосомная теория наследственности
Хромосомная теории наследственности экспериментально обоснована Т. Морганом с сотрудниками в 1911-1926 гг.
♦ Основные положения хромосомной теории наследственности:
■ гены, ответственные за хранение и передачу наследственной информации, локализованы в определенных участках (локусах) хромосом; различные хромосомы имеют разное число генов;
■ аллельные гены занимают одинаковые локусы в гомологичных хромосомах;
■ в хромосомах гены располагаются в линейной последовательности (друг за другом) и не перекрываются;
■ гены гомологичных хромосом образуют группу сцепления и наследуются преимущественно вместе; число групп сцепления равно гаплоидному набору хромосом (т.е. числу пар гомологичных хромосом);
■ между гомологичными хромосомами возможен обмен аллельными генами (кроссинговер);
■ вероятность кроссинговера пропорциональна расстоянию между генами в хромосомах.
Генетические карты
Генетическая карта хромосом — схема, на которой отображается взаимное расположение генов, находящихся в одной группе сцепления, с учетом их порядка следования и относительных расстояний между ними.
■ Возможность картирования хромосом основана на постоянстве процента кроссинговера между определенными генами.
Расстояние между генами выражается в морганидах.
■ Одна морганида — такое расстояние между генами, при котором вероятность кроссинговера равна 1 %.
♦ Значение генетических карт:
■ в селекции они позволяют прогнозировать возможность получения организмов с определенными сочетаниями признаков;
■ являются основой для генной инженерии;
■ в медицине используются для диагностики ряда тяжелых наследственных заболеваний человека;
■ сравнение генетических карт разных видов живых организмов помогает установить особенности эволюционного процесса.
Цитоплазматическая наследственность
Цитоплазматическая наследственность — наследственность, связанная с действием генов, находящихся в органоидах цитоплазмы, содержащих ДНК (митохондриях и пластидах).
■ Такие гены способны к автономной репликации и равномерному распределению между дочерними клетками.
■ В передаче признаков цитоплазматическая наследственность имеет второстепенное значение.
■ Цитоплазматическое наследование осуществляется только через материнский организм (в мужских половых клетках цитоплазмы мало и в ней митохондрии и пластиды отсутствуют). Примеры: с мутациями генов, локализованных в митохондриях, связано наследование нарушений в действии дыхательных ферментов у дрожжей; с мутациями генов, локализованных в пластидах, связано, в частности, наследование пестролистности у ряда растений (ночной красавицы, львиного зева и др.).
Читайте также: