
Хромосома как группа сцепления генов хромосомная теория наследования т моргана кариотип кариограмма
Обновлено: 25.06.2024
1. Хромосомная теория наследственности. Законы сцепления признаков
2. Регламент
3. Актуальность
4. Третий закон Менделя
Как уже говорилось на первом уроке, для доказательства
третьего закона, Мендель скрестил горох, отличающийся по
двум парам альтернативных признаков (желтый и гладкий,
зеленый и морщинистые) и в результате получил расщепление
во втором поколении 9 : 3 : 3 : 1.
При этом он доказал, что гены, отвечающие за формирование
исследуемых признаков, расположены в разных хромосомах, и
при образовании гамет у гибридов первого поколения они
будут комбинироваться независимо друг от друга. Поэтому он
и получил четыре типа гамет AB, Ab, aB, ab.
При этом если подсчитать расщепление, то дигибридное
скрещивание (которое лежит в основе третьего закона) ничто
иное, как два независимо идущих моногибридных
скрещивания, результаты которых как бы накладываются друг
на друга.
5. Третий закон Менделя
Полученные результаты дигибридных скрещиваний
позволили Менделю сформулировать закон
независимого наследования.
При скрещивании двух гомозиготных особей,
отличающихся по двум и более парам
альтернативных признаков, гены и
соответствующие им признаки передаются
потомству независимо друг от друга и
комбинируются во всех возможных
сочетаниях.
Таким образом, вероятность
сочетания аллелей в генотипе
равны:
1.
Желтые и гладкие - 9/16
2.
Желтые и морщинистые
– 3/16
3.
Зеленые гладкие – 3/16
4.
Зеленые морщинистые –
1/16
Соотношение фенотипов во
втором поколении составило
9:3:3:1
7. Краткое изложение гипотезы Менделя
1.
2.
3.
4.
5.
6.
Каждый признак данного организма контролируется парой аллелей.
Если организм содержит два различных аллеля по данному
признаку, то один из них (доминантный) может проявляться,
полностью подавляя проявление другого признака (рецессивного).
При мейозе каждая пара аллелей разделяется (расщепляется) и
каждая гамета получает по одному от каждой пары аллелей (закон
расщепления).
При образовании мужских и женских гамет в каждую из них может
попасть любой аллель из одной пары вместе с любым другим из
другой пары (закон независимого распределения).
Каждый аллель передается из поколения в поколение как
дискретная неизменяющаяся единица.
Каждый организм наследует по одному аллелю (для каждого
признака) от каждой из родительских особей.
8. Краткое изложение гипотезы Менделя
Описанный механизм наследования при
дигибридном скрещивании в типичном соотношении
9 : 3 : 3 : 1 касается только тех признаков, которые
контролируются генами, локализованными в разных
хромосомах. Если же гены находятся в одной и той
же хромосоме, то независимого распределения
такого типа наблюдается не во всех случаях.
9. Хромосомная теория наследственности
В середине XIX в., когда Мендель проводил свои
эксперименты и формулировал закономерности, имеющие
всеобщее и фундаментальное значение для генетики и
биологии в целом, научных знаний было недостаточно для
понимания механизмов наследования, и ученые не смогли
оценить значения его открытий.
К концу XIX – началу ХХ вв. благодаря существенному
улучшению оптических характеристик микроскопов, были
открыты митоз и мейоз, а в 1900 г. заново переоткрыты
законы Менделя почти одновременно тремя учеными – де
Фризом, Корренсом и Черкмаком, которые оценили их
должным образом.
10. Хромосомная теория наследственности
Американские
ученые У. Сэттон и
Т. Бовери высказали
предположение, что
хромосомы служат
носителями
менделевских
фактором, и
сформулировали
хромосомную
теорию
наследственности.
11. Хромосомная теория наследственности
Согласно этой теории, каждая пара факторов (генов)
локализована в паре гомологичных хромосом, причем каждая
хромосома несет по одному из этих факторов.
Поскольку число признаков у любого организма во много раз
больше числа его хромосом , видимых под микроскопом,
каждая хромосома должна содержать множество факторов.
Дальнейшее обоснование хромосомной теории наследования
принадлежит американскому генетику Т. Моргану.
12. Работа Т. Моргана
Морган установил, что гены, находящиеся в одной и той же
хромосоме, называются сцепленными, а все гены одной
хромосомы образуют группу сцепления и наследуются
совместно, т.е. сцеплено. Это явление получило название закон
Моргана.
Для своего исследования Морган выбрал муху дрозофилу. При
скрещивании мухи с серым телом и длинными крыльями с
мухой с черным телом и зачаточными крыльями, он получил
помимо мух с родительскими фенотипами еще и особей с
новыми сочетаниями признаков. Эти новые фенотипы
называют рекомбинантными.
Число групп сцепления =
гаплоидному набору хромосом
– у женщин 23, у мужчин 24
13. Работа Т. Моргана
В результате опыта Морган получил
следующие результаты:
41,5% - серое тело, длинные крылья
41,5% - черное тело, зачаточные
крылья
8,5% серое тело, зачаточные крылья
8,5% черное тело, длинные крылья
14. Работа Т. Моргана
На основе этих результатов Морган постулировал, что:
1.
Изучаемые гены локализованы в хромосомах
2.
Оба гена находятся сцеплено, т.е. в одной хромосоме
3.
Аллели каждого гена находятся в гомологичных хромосомах
4.
Во время мейоза между гомологичными хромосомами происходит
обмен аллелями.
Появление рекомбинантных сочетаний аллелей у 17% потомков было
объяснено на основе пункта 4. Это явление получило название
кроссинговера.
Вероятность кроссинговера зависит
от расстояния между генами
1 сантиморганида = 1% кроссинговера,
причем, чем дальше друг от друга
находятся сцепленные гены, тем выше
частота кроссинговера!
15. Основные положения хромосомной теории наследования
1.
2.
3.
4.
5.
6.
Гены локализованы в хромосомах. При этом различные
хромосомы содержат неодинаковое число генов. Кроме того,
набор генов каждой из негомологичных хромосом уникален
Аллельные гены занимают одинаковые локусы в
гомологичных хромосомах
Гены расположены в хромосоме в линейной
последовательности
Гены одной хромосомы образуют группу сцепления, то есть
наследуются преимущественно сцепленно. Число групп
сцепления равно гаплоидному числу хромосом данного вида
Сцепление нарушается в результате кроссинговера, частота
которого прямо пропорциональна расстоянию между генами в
хромосоме
Каждый биологический вид характеризуется определенным
набором хромосом —кариотипом.
16. Линейное расположение генов Генетические карты
Один из ученых Моргана, А.Х. Стергевант высказал, что
частота рекомбинаций свидетельствует о линейном
расположении генов вдоль хромосомы и, что эта частота
отражает относительное расположение генов в хромосоме, чем
дальше друг от друга находятся сцепленные гены, тем выше
частота кроссинговера.
Данные о частоте рекомбинаций важны прежде всего потому,
что они дают генетикам возможность составлять карты
относительного расположения генов в хромосомах. Кары
строятся путем прямого перевода частоты кроссинговера в
предполагаемые расстояния на хромосоме.
17. Линейное расположение генов Генетические карты
На практике обычно определяют частоту кроссинговера
по меньшей мере для трех генов одновременно; этот
метод, называемый триангуляцией, позволяет определить
не только расстояние между генами, но и их
последовательность.
При построении карт могут возникать затруднения,
создаваемые двойным кроссинговером: особенно, когда
гены разделены большим расстоянием, так как число
кроссоверов бывает при этом меньше их фактического
числа.
19. Пример решения задач на сцепленное наследование
Задача
У человека развитие дальтонизма и
гемофилии обусловлено действием
рецессивных генов, локализованных
в Х-хромосоме на расстоянии 9,8%
сантиморганид друг от друга.
Здоровая женщина, отец которой
страдал гемофилией, вступает в брак
со здоровым мужчиной. Известно,
что мать женщины была
дальтоником, но среди ее
родственников гемофилии не было.
1.
Какой процент гамет женщины
содержат одновременно гены
гемофилии и дальтонизма?
2.
Сколько разных генотипов может
быть среди детей этой
супружеской пары?
Дано
Механизм наследования сцепленных генов, а также местоположение некоторых сцепленных генов установил американский генетик и эмбриолог Т. Морган. Он показал, что закон независимого наследования, сформулированный Менделем, действителен только в тех случаях, когда гены, несущие независимые признаки, локализованы в разных негомологичных хромосомах. Если же гены находятся в одной и той же хромосоме, то наследование признаков происходит совместно, т. е. сцепленно. Это явление стали называть сцепленным наследованием, а также законом сцепления или законом Моргана.
Закон сцепления гласит: сцепленные гены, расположеные в одной хромосоме, наследуются совместно (сцепленно).Группа сцепления — все гены одной хромосомы. Число групп сцепления равно количеству хромосом в гаплоидном наборе. Например, у человека 46 хромосом — 23 группы сцепления, у гороха 14 хромосом — 7 групп сцепления, у плодовой мушки дрозофилы 8 хромосом — 4 группы сцепления.Неполное сцепление генов — результат кроссинговера между сцепленными генами, поэтому полное сцепление генов возможно у организмов, в клетках которых кроссинговер в норме не происходит.
ХРОМОСОМНАЯ ТЕОРИЯ МОРГАНА. ОСНОВНЫЕ ПОЛОЖЕНИЯ.
Результатом исследований Т. Моргана стало создание им хромосомной теории наследственности:
1)гены располагаются в хромосомах; различные хромосомы содержат неодинаковое число генов; набор генов каждой из негомологичных хромосом уникален;
2)каждый ген имеет определенное место (локус) в хромосоме; в идентичных локусах гомологичных хромосом находятся аллельные гены;
3)гены расположены в хромосомах в определенной линейной последовательности;
4)гены, локализованные в одной хромосоме, наследуются совместно, образуя группу сцепления; число групп сцепления равно гаплоидному набору хромосом и постоянно для каждого вида организмов;
5)сцепление генов может нарушаться в процессе кроссинговера, что приводит к образованию рекомбинантных хромосом; частота кроссинговера зависит от расстояния между генами: чем больше расстояние, тем больше величина кроссинговера;
6)каждый вид имеет характерный только для него набор хромосом — кариотип.
Полное и неполное сцепление генов.
Гены в хромосомах имеют разную силу сцепления. Сцепление генов может быть: полным, если между генами, относящимися к одной группе сцепления, рекомбинация невозможна и неполным, если между генами, относящимися к одной группе сцепления, возможна рекомбинация.
Генетические карты хромосом.
Это схемы относительного расположения сцепленных между собой
наследственных факторов — генов. Г. к. х. отображают реально
85. Хромосомный механизм наследования пола. Цитогенетические методы определения пола.
Пол характеризуется комплексом признаков, определяемых генами, расположенными в хромосомах. У видов с раздельнополыми особями хромосомный комплекс самцов и самок неодинаков, цитологически они отличаются по одной паре хромосом, ее назвали половыми хромосомами. Одинаковые хромосомы этой пары назвали X(икс)- хромосомами. Непарную, отсутствующую у другого пола- Y (игрек)- хромосомой; остальные, по которым нет различий аутосомами (А). У человека 23 пары хромосом. Из них 22 пары аутосом и 1 пара половых хромосом. Пол с одинаковыми хромосомами XX, образующий один тип гамет (с X- хромосомой), называют гомогаметным,другой пол, с разными хромосомами XY, образующий два типа гамет (с X-хромосомой и с Y-хромосомой), - гетерогаметным. У человека, млекопитающих и других организмов гетерогаметный пол мужской; у птиц, бабочек - женский.
X- хромосомы, помимо генов, определяющих женский пол, содержат гены, не имеющие отношения к полу. Признаки, определяемые хромосомами, называются признаками, сцепленными с полом. У человека такими признаками являются дальтонизм (цветная слепота) и гемофилия (несвертываемость крови). Эти аномалии рецессивны, у женщин такие признаки не проявляются, если даже эти гены несет одна из X- хромосом; такая женщина является носительницей и передает их с Х — хромосомой своим сыновьям.
Цитогенетический метод определения пола. Он основан на микроскопическом изучении хромосом в клетках человека. Применение цито генетического метода позволяет не только изучать нормальную морфологию хромосом и кариотипа в целом, определять генетический пол организма, но, главное, диагностировать различные хромосомные болезни, связанные с изменением числа хромосом или с нарушением их структуры. В качестве экспресс- метода, выявляющего изменение числа половых хромосом, используют метод определения полового хроматинав неделящихся клетках слизистой оболочки щеки. Половой хроматин, или тельце Барра, образуется в клетках женского организма одной из двух Х- хромосом. При увеличении количества Х — хромосом в кариотипе организма в его клетках образуются тельца Барра в количестве на единицу меньше числа хромосом. При уменьшении числа хромосом тельце отсутствует. В мужском кариотипе Y- хромосома может быть обнаружена по более интенсивной люмисценции по сравнению с другими хромосомами при обработке их акрихинипритом и изучении в ультрафиолетовом свете.
Особенности строения хромосом. Уровни организации наследственного материала. Гетеро- и эухроматин.
Морфология хромосом
При микроскопическом анализе хромосом, прежде всего, видны различия их по форме и величине. Строение каждой хромосомы сугубо индивидуальное. Можно заметить также, что хромосомы обладают общими морфологическими признаками. Они состоят из двух нитей — хроматид, расположенных параллельно и соединенных между собой в одной точке, названной центромерой или первичной перетяжкой. На некоторых хромосомах можно видеть и вторичную перетяжку. Она является характерным признаком, позволяющим идентифицировать отдельные хромосомы в клетке. Если вторичная перетяжка расположена близко к концу хромосомы, то дистальный участок, ограниченный ею, называют спутником. Хромосомы, содержащие спутник, обозначаются как АТ-хромосомы. На некоторых из них в телофазе происходит образование ядрышек.
Концевые участки хромосом имеют особую структуру и называются теломерами. Теломерные районы обладают определенной полярностью, препятствующей их соединению друг с другом при разрывах или со свободными концами хромосом.
Участок хроматиды (хромосомы) от теломеры до центромеры называют плечом хромосомы. Каждая хромосома имеет два плеча. В зависимости от соотношения длин плеч выделяют три типа хромосом: 1) метацентрические (равноплечие); 2) субметацентрические (неравноплечие); 3) акроцентрические, у которых одно плечо очень короткое и не всегда четко различимо. (р — короткое плечо, q — длинное плечо). Изучение химической организации хромосом эукариотических клеток показало, что они состоят в основном из ДНК и белков: гистонов и протомите (в половых клетках), которые образуют нуклеопротеиновый комплекс—хроматин, получивший свое название за способность окрашиваться основными красителями. Белки составляют значительную часть вещества хромосом. На их долю приходится около 65% массы этих структур. Все хромосомные белки разделяются на две группы: гистоны и негистоновые белки.
Гистоны представлены пятью фракциями: HI, Н2А, Н2В, НЗ, Н4. Являясь положительно заряженными основными белками, они достаточно прочно соединяются с молекулами ДНК, чем препятствуют считыванию заключенной в ней биологической информации. В этом состоит их регуляторная роль. Кроме того, эти белки выполняют структурную функцию, обеспечивая пространственную организацию ДНК в хромосомах.
Число фракций негистоновых белков превышает 100. Среди них ферменты синтеза и процессинга РНК, редупликации и репарации ДНК. Кислые белки хромосом выполняют также структурную и регуляторную роль. Помимо ДНК и белков в составе хромосом обнаруживаются также РНК, липиды, полисахариды, ионы металлов.

В видеоуроке рассматривается сцепленное наследование признаков, закон Моргана, а также генетическое определение пола. В данном уроке приводятся следующие понятия: кроссинговер, хромосомная карта, неполное сцепление, полное сцепление, аутосомы.
В данный момент вы не можете посмотреть или раздать видеоурок ученикам
Чтобы получить доступ к этому и другим видеоурокам комплекта, вам нужно добавить его в личный кабинет, приобрев в каталоге.
Получите невероятные возможности



Конспект урока "Сцепленное наследование. Закон Моргана. Генетика пола"
Третий закон Менделя — закон независимого наследования признаков, или независимого комбинирования генов, ― справедлив в тех случаях, когда гены рассматриваемых признаков располагаются в разных гомологичных хромосомах.
Что же будет, если признаки будут располагаться в одной хромосоме?
После открытых Менделем законов в 1906 году учёные Уильям Бэтсон и Реджинальд Пэннет обнаружили, что в некоторых случаях расщепления по правилам Менделя не происходит.
Например, у душистого горошка два признака — форма пыльцы и окраска цветков — не дают независимого расщепления в потомстве в соотношении 3:1 (три к одному) и потомки остаются похожи на родительскую особь. При последующем анализе оказалось, что гены этих двух признаков лежат в одной хромосоме.
Изучением наследования признаков, гены которых локализованы в одной хромосоме, занимался американский генетик Томас Морган.
Он предложил закон сцепленного наследования (закон Моргана): гены, которые находятся в одной хромосоме, при мейозе попадают в одну гамету, то есть наследуются сцеплено.

Что значит сцеплено? Это значит, что гены находятся в одной хромосоме. И между ними возникают силы сцепления, то есть силы взаимодействия. И чем ближе эти гены, тем сильнее взаимодействие.
Каковы же принципы наследования генов, расположенных в одной хромосоме?
Вспомним что Мендель проводил свои опыты на горохе. При дигибридном скрещивании во втором поколении соотношение между жёлтыми и зелёными горошинами составляло 3:1.
У Моргана основными объектами для изучения были фруктовые мушки дрозофилы. Дрозофила имеет диплоидный набор хромосом равный 8.
Морган скрещивал двух мушек: самка была серой с длинными крыльями, а самец с черным телом и зачаточными крыльями. В первом поколении особь имела серое тело и длинные крылья.
Значит, ген, определяющий серую окраску брюшка, доминирует над темной окраской, а ген, обуславливающий развитие нормальных крыльев, — над геном недоразвитых (зачаточных) крыльев.
При дальнейшем скрещивании между собой гибридных мух первого поколения во втором поколении появляются два фенотипа вместо четырёх ожидаемых.
Это позволило сделать вывод, что гены окраски тела и длины крыльев дрозофил находятся в одной хромосоме.
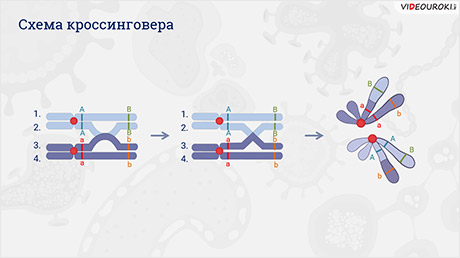
Вспомним, что символы А-большое Бэ-большое А-большое Бэ-большое и а-малое бэ-малое а-малое бэ-малое располагаются как бы друг под другом с двумя чёрточками. Чёрточками условно обозначают хромосомы.
Однако в этих же опытах было описано отклонение от закона Моргана.
При скрещивании дигетерозиготной особи, которая получилась в первом поколении, и чёрной мушки с зачаточными крыльями, имеющей оба рецессивных признака. Получился необычный результат: среди гибридов второго поколения было небольшое число особей с перекомбинацией тех признаков, гены которых лежат в одной хромосоме.
В потомстве явно преобладали особи с признаками родительских форм (41,5 % были серые длиннокрылые и 41,5 % — черные с зачаточными крыльями), и лишь незначительная часть мушек имела иное, чем у родителей, сочетание признаков (8,5 % были серые с зачаточными крыльями и 8,5 % — черные длиннокрылые).
Что-бы объяснить появление особей с рекомбинантными признаками, необходимо вспомнить деление мейоза. А именно то, что в профазу первого мейоза гомологичные хромосомы конъюгируют (сближаются) и могут обмениваться гомологичными участками. Этот процесс называется кроссинговером. Он необходим для повышения разнообразия потомков.
Процесс кроссинговера был открыт Томасом Морганом и его сотрудниками при очередном скрещивании дрозофил.
При кроссинговере происходит нарушение закона Моргана, и гены одной хромосомы не наследуются сцеплено, так как часть из них заменяется на аллельные гены гомологичной хромосомы.
Происходит обмен участками хромосом между генами А и В, появляются гаметы А-большое бэ-малое и а-малое Бэ-большое, и, как следствие, в потомстве образуются четыре группы фенотипов.
Явление кроссинговера позволило лаборатории Томаса Моргана установить расположение каждого гена в хромосоме и составить хромосомные карты.
Хромосомная карта — это графическое изображение хромосомы, на котором определённые локусы (гены) отмечены соответственно расстоянию между ними.
Определить, в каких участках хромосом произойдёт кроссинговер, достаточно сложно.
Томас Морган доказал, что частота кроссинговера между генами прямо пропорциональна расстоянию между ними в хромосоме. Другими словами, можно сказать, что чем дальше гены находятся друг от друга в хромосоме, тем чаще между ними происходит кроссинговер.
Например, если рассматривать 2 гена А и Дэ, мы можем увидеть 2 случая.
Если два гена А и Дэ находятся по разные стороны перекрёста, то они разойдутся в разные хромосомы при любом варианте кроссинговера. В данном случае А-большое дэ-малое и а-малое Дэ-большое.
Но если гены, например, А-большое и Дэ-большое, расположены в хромосоме рядом по одну сторону от перекрёста, тогда после прохождения кроссинговера новых сочетаний аллелей данных двух генов мы не увидим.
Различают неполное и полное сцепление. Неполное сцепление — это разновидность сцепленного наследования, при котором гены анализируемых признаков располагаются на некотором расстоянии друг от друга, что делает возможным кроссинговер между ними.

Полное сцепление — это разновидность сцепленного наследования, при котором гены анализируемых признаков располагаются так близко друг к другу, что кроссинговер между ними становится невозможным.
Исходя из своих открытий, Томас Морган сформулировал хромосомную теорию наследственности.
Первое положение этой теории гласит: ген представляет собой участок хромосомы. Хромосомы, таким образом, представляют собой группу сцепления генов.
Второе положение утверждает, что аллельные гены (гены, которые отвечают за один признак) расположены в строго определённых местах (локусах) гомологичных хромосом.
Согласно третьему положению, гены располагаются в хромосомах линейно, то есть друг за другом.
В ещё одном дополнительном положении говорится, что в процессе образования гамет между гомологичными хромосомами происходит конъюгация, в результате которой они могут обмениваться аллельными генами, т. е. может происходить кроссинговер.
Генетическое определение пола
Большинство животных являются раздельнополыми организмами. Но как так получается, что на свет особи женского и мужского пола рождаются примерно в одинаковом количестве? То есть расщепление по половой принадлежности происходит в соотношении 1:1 (один к одному).
Ещё Грегор Мендель, изучая закономерности наследования признаков, заметил, что расщепление 1:1 в потомстве по какому-либо признаку наблюдается в тех случаях, когда одна из родительских особей гетерозиготная (А-большое а-малое) по этому признаку, а вторая — рецессивная гомозиготная (а-малое а-малое).
В результате наблюдений было сделано предположение, что один из полов (тогда было неясно, какой именно) гетерозиготен, а второй гомозиготен по гену, который определяет пол организма.
Понять, как определяется пол будущей особи, учёные смогли в начале 20-го века. Современную теорию наследования пола разработал Томас Морган и его сотрудники. Они установили, что самцы и самки различаются по набору хромосом.
Кариотип каждого организма содержит хромосомы, одинаковые у обоих полов, их называют — аутосомами. Аутосомы — это не половые хромосомы.
А хромосомы, по которым женский и мужской пол отличаются друг от друга, называются — половыми хромосомами.
В хромосомном наборе клеток млекопитающих и человека женский пол имеет две одинаковые хромосомы (тип X (икс) X (икс)), а мужской пол - неодинаковые половые хромосомы (тип XУ, где X-хромосома более крупная).
Томас Морган заметил, что у самки дрозофилы три пары аутосом и одна пара одинаковых, так называемых Икс-хромосом. А у самцов те же три пары аутосом и две разные половые хромосомы — икс-игрек.
При формированиии половых клеток — гамет — во время мейоза у самок будет образовываться один тип гамет: 3 аутосомы плюс Икс-хромосома. А у самцов в одинаковом количестве будут образовываться два типа гамет: 3 аутосомы + Икс-хромосома или 3 аутосомы плюс Игрек-хромосома.
Теоретически при оплодотворении гамета самки с равной вероятностью может встретиться с любой из гамет самца.
Например, если при оплодотворении с яйцеклеткой сольётся сперматозоид с икс-хромосомой, то из зиготы разовьётся самка. А если с игрек-хромосомой, то самец.
Соотношение полов при таком скрещивании теоретически всегда будет 1:1.
Таким образом пол будущей особи определяется во время оплодотворения и зависит от того, какой набор половых хромосом сформируется в тот момент.
Так как самки дрозофилы способны производить только один тип гамет (с икс (Х) половой хромосомой), их пол называют гомогаметным.
А так как самцы дрозофилы производят два вида гамет (и с Икс (Х) и с Игрек (Y) половыми хромосомами, их пол называют гетерогаметным.
Хромосомный набор человека представлен 46 хромосомами. Из них у женщины и мужчины в каждой клетке по 22 пары аутосом-неполовых хромосом. И по паре половых хромосом. У женщины это две икс-хромосомы. У мужчин одна пара половых из которых одна икс, вторая игрек.
Наследование признаков, сцепленных с полом.
В Y-хромосоме находятся гены, которые необходимы для нормального развития мужских половых признаков, а в Икс-женских.
Но не все эти гены отвечают за признаки, которые имеют отношение к полу.
Расположение гена в половой хромосоме называют сцеплением гена с полом.
В соматических клетках женщины по две х-хромосомы, поэтому за каждый признак отвечает по два расположенных в них гена, а в клетках организма мужчины всего одна х-хромосома и все полторы сотни генов, расположенные в ней (и доминантные, и рецессивные), ― обязательно проявляются в фенотипе.
Изучением наследования генов, локализованных в половых хромосомах, занимался Томас Морган.
Он скрещивал красноглазых самок с белоглазыми самцами. Так как у дрозофилы красный цвет глаз доминирует над белым, то в первом поколении все потомство оказывалось красноглазым.
Далее при скрещивании между собой гибридов первого поколения во втором поколении все самки оказываются красноглазыми, а среди самцов — половина белоглазых и половина красноглазых.
При скрещивании между собой белоглазых самок и красноглазых самцов в первом поколении все самки оказываются красноглазыми, а самцы белоглазыми. Во втором поколении половина самок и самцов — красноглазые, половина — белоглазые.
Объяснить полученные результаты наблюдаемого расщепления по окраске глаз Морган смог, только предположив, что ген, отвечающий за окраску глаз, локализован в Х-хромосоме.
Икс с доминантным геном отвечает за красный цвет глас. Икс с рецессивным геном за белый цвет глаз, а Y-хромосома таких генов не содержит.
У человека некоторые заболевания сцеплены с полом. К ним относится, например, гемофилия и дальтонизм.
Рассмотрим наследование дальтонизма.

Нормальное цветовосприятие обусловлено доминантным аллелем, который находится в X-хромосоме. Его рецессивная аллельная пара в гомо- и гетерозиготном состоянии приводит к развитию дальтонизма.
Отсюда понятно, почему дальтонизм чаще встречается у мужчин, чем у женщин: у мужчин только одна X-хромосома, и если в ней находится рецессивный аллель, определяющий дальтонизм, он обязательно проявляется.
У женщины две X хромосомы: она может быть, как гетерозиготной, так и гомозиготной по этому гену. И только при гомозиготном сочетании этих генов дальтонизм проявится.
Одной из самых важных вех в развитии генетики стали экспериментальные работы Т. Моргана (1911-1914 гг.) и его сотрудников (А. Стертеван, К. Бриджис, Г. Меллер) с плодовой мушкой дрозофилой. Ими была установлена связь генов с хромосомами. Эти работы экспериментально доказали хромосомную теорию наследственности.
1901-1902 гг. – Мак-Кланг предположил, что хромосомы являются определяющим пола (1905 г –Вильсон и Стивенс подтвердили половые хромосомы).
1902 г - Бовери и Сэттон обнаружили, что количество признаков намного превосходит количество хромосом.
1902 г – Сэттон установил сходство поведения хромосом во время образования гамет, оплодотворения и передачи менделеевских наследственных факторов.
Было выдвинуто предположение: в каждой хромосоме должно быть множество различных задатков, которые должны наследоваться совместно.
1902 – 1907 гг. – Бовери проводил эксперименты над яйцами морских ежей и доказал, что требуется наличие всех хромосом, присущих непосредственно виду. Так же описаны все хромосомы, относящиеся к определенному полу
1905 г – Стивенсон установил различия в половом наборе самок и самцов насекомых.
1902-1903 гг. – Сэттон и Бовери сформировали основные положения хромосомной теории наследственности:
1. Хромосома – материальный носитель наследственных задатков;
2. Пара наследственных задатков (отца и матери) локализованы в паре хромосом отцовского и материнского происхождения, при этом каждая хромосома несет по одному наследственному задатку из пары;
3. Каждая хромосома содержит множество наследственных задатков;
4. Наследственные задатки, расположенные в одной паре, наследуются совместно.
б) Экспериментальное доказательство хромосомной теории наследственности (работы Т. Моргана)
1911-1914 гг – Томас Морган в экспериментальном исследовании с мушкой дрозофилы экспериментально доказал основные положения хромосомной теории (в 1933 г – нобелевская премия).
Разработана теория линейного расположения гена в хромосоме и вытекающие от сюда процессы сцепления и кроссинговер.
В качестве исследования использовал дрозофилу melanogaster, так как:
1. Маленький размер
3. Легко разводится на искусственной питательной среде
4. Обладает высокой плодовитостью (1 особь – 100 потомков)
5. Имеет ряд менделирующих альтернативных признаков
6. Короткий цикл индивидуального развития (10 дней)
7. Минимальное количество хромосом (4 пары)
8. Самец имеет полное сцепление
Красные глаза доминируют над белыми. Морган взял красноглазую самку и белоглазого самца, скрестил их и в первом поколении наблюдал единообразие (красные глаза), что подтверждает первый закон Менделя.
При скрещивании гибридов первого поколения он наблюдал расщепление по фенотипу 3:1, по генотипу 1:2:1, что доказывало второй закон Менделя.
Затем скрестил красноглазого самца и белоглазую самку, в первом поколении наблюдал расщепление 1:1. Установил закономерность, что красноглазыми были самки, а белоглазыми – самцы. Сделал вывод, что признак, отвечающий за цвет глаз, находится в хромосоме, преимущественно в Х и наследуется совместно и не подчиняется законам Менделя.
3.1. Взял альтернативные признаки: цвет тела (черный, серый), форму крыльев (длинные, редуцированные), чистые линии (не дают расщепления в потомстве). Серый цвет тела доминирует над черным. Длинные крылья доминируют над короткими.
Скрещивал гомозиготную самку по обоим доминантным признакам и гомозиготного самца по обоим рецессивным признакам. Получил единообразие первого поколение поколения (серый цвет, длинные крылья).
Затем провел анализирующее скрещивание (самец F1 с гомозиготной самкой). В результате получил расщепление 1:1, полностью соответствуют родительским. А Мендель получил бы 4 фенотипических класса в равном соотношении 25% каждый (1:1:1:1).
3.2. Скрестил самку гибридов F1 с гомозиготным самцом и получил иные результаты: 4 фенотипических класса, с соотношением: 2 класса с признаками родителей (большинство) – 83% (по 41,5%) и 2 с перекомбинацией генов – 17% (по 8,5%).
При скрещивании гибридов первого поколения, самки – доминантные, самцы – рецессивные, 4 фенотипических класса 83% с признаками родителей и 17% с перекомбинацией признаков.

Это позволило Т. Моргану и его коллегам сформулировать основные положения хромосомной теории наследственности:
1. Гены локализованы в хромосомах. При этом различные хромосомы содержат неодинаковое число генов. Кроме того, набор генов каждой из негомологичных хромосом уникален.
2. Аллельные гены занимают одинаковые локусы в гомологичных хромосомах.
3. Гены расположены в хромосоме в линейной последовательности.
4. Гены одной хромосомы образуют группу сцепления, то есть наследуются преимущественно сцеплено, благодаря чему происходит сцепленное наследование некоторых признаков. Число групп сцепления равно гаплоидному числу хромосом данного вида (у гомогаметного пола) или больше на 1 (у гетерогаметного пола).
5. Сцепление нарушается в результате кроссинговера, частота которого прямо пропорциональна расстоянию между генами в хромосоме (поэтому сила сцепления находится в обратной зависимости от расстояния между генами).
6. Каждый биологический вид характеризуется определенным набором хромосом -- кариотипом.
в) Сцепление генов (полное, неполное). Группы сцепления у разных биологических видов.
Сцепление генов – гены находятся в одной хромосоме.
Группа сцепления - совокупность генов, локализованных в одной хромосоме. Число групп сцепления равно числу пар хромосом в диплоидном наборе данного вида.
Сцепление неаллельных генов может быть полным (если гены всегда передаются вместе) и неполным (встречается чаще):
Полное сцепление – разновидность сцепленного пола, при котором гены анализируемых признаков располагаются близко друг к другу
Неполное сцепление – это такое, при котором анализируемое скрещивание гетерозиготного организма по двум признакам с рецессивной формой, мы получаем не два фенотипа, а четыре.
Неполное сцепление генов обусловлено кроссинговером, который происходит в профазе (стадия пахитены) первого деления митоза. Кроссинговер - это процесс обмена одинаковыми участками между гомологичными хромосомами. В результате кроссинговера возникают новые комбинации генов в гомологичных хромосомах. Генетические рекомбинации приводят к формированию в гаметогенезе различных типов гамет: одни гаметы с хромосомами, которые не подвергались кроссинговеру (некроссоверные гаметы), другие гаметы с хромосомами, которые участвовали в кроссинговере (кроссоверные гаметы).
Вероятность кроссинговера и количество кроссоверных (рекомбинантных) гамет (особей) зависит от расстояния между генами (прямая зависимость). Чем ближе друг к другу расположены гены в хромосоме, тем реже происходит их расхождение при кроссинговере, и, наоборот, чем дальше друг от друга отстоят гены, тем чаще возможно нарушение их сцепления. Мерой расстояния между генами является частота кроссинговера, которая определяется через отношение числа кроссоверных особей в потомстве (образовались при участии кроссоверных гамет) к общему числу потомков в данном скрещивании, выраженное в процентах. Единицей расстояния между генами является морганида. 1 морганида соответствует 1% кроссинговера (или 1%кроссоверных особей, или 1% кроссоверных гамет).
Следует отметить, что у подавляющего большинства животных, растений и человека кроссинговер происходит в одинаковых долях как у особей женского, так и мужского пола. К исключениям относятся, лишь самец дрозофилы и самка тутового шелкопряда, у которых кроссинговер не происходит, и образуются только некроссоверные гаметы.
Группы сцепления – гены, расположенные в линейной последовательности в одной хромосоме и наследуемые совместно.
Неаллельные гены, локализованные в одной хромосоме образуют группу сцепления. Число групп сцепления равно числу пар хромосом в диплоидном наборе данного вида. Например, у самки дрозофилы имеется восемь хромосом и соответственно четыре группы сцепления; у самца дрозофилы имеется также восемь хромосом, но групп сцепления будет пять: три за счет трех пар аутосом, одна за счет Х-хромосомы и одна за счет Y-хромосомы. Выделение отдельных групп сцепления у самца по Х- и Y-хромосомам определяется тем, что эти хромосомы не являются гомологичными и имеют существенные различия по генному составу. Аналогичным образом у человека, женский организм имеет 23 группы сцепления, а мужской — 24 группы сцепления.
г) Основные положения хромосомной теории наследственности.
1. Гены локализуются в хромосоме в линейной последовательности. Каждый ген занимает определённое место – локус;
2. Гены, локализованные в одной хромосоме, наследуются совместно, образуя группу сцепления. Число групп сцепления равно гаплоидному набору хромосом;
3. Сила сцепления генов в хромосоме зависит от расстояния между ними. Чем ближе расположены гены друг к другу, тем сильнее сила их сцепления;
4. Сцепление генов может нарушаться процессом кроссинговера, в результате которого образуются рекомбинантные хромосомы. Чем сильнее сцеплены гены, тем меньше величина кроссинговера;
5. Сцепление генов и их рекомбинация в результате кроссинговера – это закономерные биологические явления, в которых выражается единство наследственности и изменчивости как свойства живого.
д) Принципы построения генетических карт. Цитологические карты хромосом.
Генетическое картирование – это определение положения какого-либо гена по отношению к другим генам в данной группе. Результат – составление генетических карт хромосом.
Генетическая карта – это отрезок прямой, на котором обозначен порядок взаимного расположения генов в хромосоме и указание расстояния между ними, которое измеряется в морганидах, или в процентах кроссинговера.
В основе – линейный порядок расположения генов. Строятся на результатах анализирующих скрещиваний.
Физический метод построения. При помощи электронного микроскопа или при некоторых видах электрофореза определяют межгенное расстояние.
Генетический метод. Определяют частоту рекомбинации генов, на основе чего строят генетическую карту.
Цитогенетический. Значение картирования – предотвращение и лечение наследственных заболеваний и ускорения изучения молекулярных механизмов, которые лежат в основе отклонений от нормы (нарушений).
Цитологические (цитогенетические) карты хромосом – схематическое изображение хромосом, при которых указывается привязка гена к конкретному участку хромосомы. Например, гены группы крови системы АВ0 расположены в длинном плече 9й хромосомы в 34 сегменте – обозначается: 9q34.субсимир12

Лекция 4. Хромосомная теория наследственности. Хромосомные карты человека.
Хромосомная теория наследственности. Хромосомные карты человека.
Хромосомная теория Т.Моргана.
Сцепленные гены, кроссинговер.
Карты хромосом человека.
Хромосомная теория Т.Моргана.
Наблюдая за большим количеством мух, Т. Морган выявил много мутаций, которые были связаны с изменением разных признаков: окраски глаз, формы крыльев, окраски тела и т.д.
При изучении наследования этих мутаций оказалось, что многие из них наследуются, сцепленно с полом.
Такие гены легко было выделить, потому что они передавались от материнских особей только потомству мужского пола, и через них - только их потомкам женского пола.
У человека признаки, наследуемые через Y-хромосому, могут быть только у лиц мужского пола, а наследуемые через Х-хромосому - у лиц как одного, так и другого пола.
При этом особь женского пола может быть гомо или гетерозиготной по генам, расположенным в Х-хромосоме, а рецессивные гены могут проявляться у нее только в гомозиготном состоянии.
У особи мужского пола только одна Х-хромосома, поэтому все локализованные в ней гены, в том числе и рецессивные, проявляются в фенотипе. Такие патологические состояния, как гемофилия (медленная свертываемость крови, обусловливающая повышенную кровоточивость), дальтонизм (аномалия зрения, при которой человек путает цвета, чаще всего красный с зеленым), наследуются у человека сцепленно с полом.
Исследование наследования, сцепленного с полом, стимулировало изучение сцепления между другими генами.
В качестве примера можно привести эксперименты на дрозофиле.
У дрозофилы существует мутация, обусловливающая черный цвет тела. Ген, ее вызывающий, рецессивен по отношению к гену серого цвета, характерному для дикого типа. Мутация, вызывающая рудиментарные крылья, также рецессивна к гену, приводящему к развитию нормальных крыльев. Серия скрещиваний показала, что ген черного цвета тела и ген рудиментарных крыльев передавались вместе, как будто оба эти признаки вызывались одним геном.
Причина такого результата заключалась в том, что гены, обусловливающие два признака, локализованы в одной хромосоме. Это явление так называемого полного сцепления генов. В каждой хромосоме расположено много генов, которые наследуются совместно, и такие гены называют группой сцепления.
Таким образом, закон независимого наследования и комбинирования признаков, установленный Г. Менделем, действует только в случае, когда гены, определяющие тот или иной признак, находятся в разных хромосомах (разных группах сцепления).
Однако гены, находящиеся в одной хромосоме, сцеплены не абсолютно.
Сцепленные гены, кроссинговер.
Причиной неполного сцепления является кроссинговер. Дело в том, что во время мейоза, при конъюгации хромосом, происходит их перекрест, и гомологичные хромосомы обмениваются гомологичными участками. Это явление называется кроссинговером. Он может произойти в любом участке гомологичных Х-хромосом, даже в нескольких местах одной пары хромосом. Причем, чем дальше друг от друга расположены локусы в одной хромосоме, тем чаще между ними следует ожидать перекрест и обмен участками.

Рисунок 17 Кроссинговер: а - схема процесса; б - варианты кроссинговера между гомологичными хромосомами
Карты хромосом человека.
В каждой группе сцепления генов содержатся сотни или даже тысячи генов.
В экспериментах А. Стертеванта в 1919 г. было показано, что гены внутри хромосомы расположены в линейном порядке.
Это было доказано путем анализа неполного сцепления в системе генов, принадлежащей к одной группе сцепления.
Изучение взаимоотношений между тремя генами при кроссинговере выявило, что в случае, если частота перекреста между генами А и В равна величине М, а между генами А и С частота обменов равна величине N, то частота перекреста между генами В и С составит М+N, или М - N, в зависимости в какой последовательности расположены гены: АВС или АСВ. И такая закономерность распространяется на все гены этой группы сцепления. Объяснение этому возможно лишь при линейном расположении генов в хромосоме.
Эти эксперименты явились основой создания генетических карт хромосом многих организмов, в том числе и человека.
Единицей генетической или хромосомной карты является сан-тиморганида (сМ). Это мера расстояния между двумя локусами, равная длине участка хромосомы, в пределах которого вероятность кроссинговера составляет 1%.
Методы изучения групп сцепления генов, такие как: генетический анализ соматических гибридных клеток, изучение морфологических вариантов и аномалий хромосом, гибридизация нуклеиновых кислот на цитологических препаратах, анализ аминокислотной последовательности белков и другие, которые позволили описать все 25 групп сцепления у человека.
Одной из основных целей исследования генома человека является построение точной и подробной карты каждой хромосомы. На генетической карте показано относительное расположение генов и других генетических маркеров на хромосоме, а также относительное расстояние между ними.
Генетическим маркером для составления карты потенциально может быть любой наследуемый признак, будь то цвет глаз или длина фрагментов ДНК. Главное при этом - наличие легко выявляемых межиндивидуальных различий рассматриваемых маркеров. Карты хромосом подобно географическим картам можно строить в разном масштабе, т.е. с разным уровнем разрешения.
Самой мелкомасштабной картой является картина дифференциального окрашивания хромосом. Максимально возможный уровень разрешения - один нуклеотид. Следовательно, самой крупномасштабной картой какой-либо хромосомы является полная последовательность нуклеотидов. Размер генома человека равен примерно 3 164,7 м.п.н.
К настоящему времени для всех хромосом человека построены мелкомасштабные генетические карты с расстоянием между соседними маркерами в 7-10 миллионов пар оснований или 7-10 Мб (мегабаз, 1Мб = 1 млн пар оснований).
Современные сведения о генетических картах человека содержат информацию о более чем 50 000 маркеров. Это означает, что они находятся в среднем на расстоянии десятков тысяч пар оснований друг от друга, и между ними расположено несколько генов.
Для многих участков, конечно же, имеются и более подробные карты, но все же большая часть генов еще не идентифицирована и не локализована.
К 2005 г. идентифицировано более 22 000 генов и около 11 000 генов картированы на отдельных хромосомах, около 6 000 генов локализованы, из них 1000 - это гены, определяющие заболевания.
Неожиданным оказалось обнаружение необычно большого числа генов на хромосоме 19 (более 1400), что превышает число генов (800), известных на самой большой хромосоме человека 1.

Рисунок 18 Патологическая анатомия хромосомы 3

Рисунок 19 Структура митохондриального генома
В митохондриальных генах отсутствуют интроны, а межгенные промежутки очень невелики. Эта небольшая молекула содержит 13 генов, кодирующих белки, и 22 гена транспортных РНК. Митохондриальная ДНК полностью секвенирована и на ней выявлены все структурные гены. Митохондриальные гены имеют гораздо большую, чем хромосомные, копийность (несколько тысяч на клетку).
Наследственные свойства крови.
Механизм наследования групп крови системы АВО и резус системы.
Причины и механизм возникновения осложнений при гемотрансфузии, связанных с неправильно подобранной донорской кровью.
Механизм наследования групп крови системы АВО и резус системы.
В одном локусе мог быть либо доминантный, либо рецессивный ген. Однако часто признак определяется не двумя, а несколькими генами.
Три или большее число генов, которые могут находиться в одном локусе (занимать одно и то же место в гомологичных хромосомах), называют множественными аллелями.
В генотипе одного индивида может быть не более двух генов из этого множества, однако в генофонде популяции соответствующий локус может быть представлен большим числом аллелей.
Примером является наследование группы крови.
Ген I A кодирует синтез в эритроцитах специфического белка агглютиногена А, ген I B - агглютиногена B, ген I О не кодирует никакого белка и является рецессивным по отношению к I A и I B ; I A и I B не доминируют относительно друг друга. Таким образом, генотип I О I О определяет группу крови 0 (первую); I A I A и I A I О - группу А (вторую); I B I B и I B I О - группу В (третью); I A I B - группу АВ (четвертую).
Если у одного из родителей группа крови 0, то (за исключением маловероятных ситуаций, требующих дополнительных обследований) у него не может родиться ребенок с группой крови АВ.
Причины и механизм возникновения осложнений при гемотрансфузии, связанных с неправильно подобранной донорской кровью.
По определению иммуногенетики группа крови это - феномен сочетания антигенов эритроцитов и антител в плазме.
Группа крови определяется сочетанием аллелей . в настоящее время известно более 30 видов аллелей детерминирующих группы крови. При гемотрансфузии учитываются те группы, которые могут вызвать осложнения. Это группы крови системы АВО, Rh-фактор, С, Kell. В донорской крови данных групп сохраняются антитела. В других известных группах антитела в донорской крови быстро разрушаются.
На рис. 20 а) показаны группы крови системы АВО, где антитела, соответствующие антигенам группы В, синего цвета, группе А – красного. Рисунок показывает, что плазма группы А имеет антитела к группе В, группы В антитела к группе А, группы АВ антител нет, группы О – антитела к группам А и В.
При гемотрансфузии (переливании крови) переливают плазму, так как, эритроциты каждого человека несут на поверхности мембраны огромное количество антигенов, специфичных для данного человека. Попав в кровь реципиента, они вызывают тяжело протекающие иммунные реакции.

Рисунок 20 Группы кови системы АВО; а) сочетание антигенов на эритроцитах и антител в плазме, b) гемолиз эритроцитов реципиента антителами донорской крови.
Если реципиенту с группой В перелить кровь (плазму) группы В, антитела в плазме немедленно вступят в взаимодействие с антигенами эритроцитов с последующим лизисом эритроцитов рис 20 b). Такой же механизм возникновения осложнений при гемотрансфузии, связанных с неправильно подобранной донорской кровью.
Практическое занятие
Решение задач, моделирующих скрещивание, сцепленное с полом наследование, наследование групп крови по системе АВО и резус систе
Читайте также: