
Генетический код менеджера типа р означает что он предпочитает осуществлять функции связанные с
Обновлено: 02.07.2024
Ранее мы подчёркивали, что нуклеотиды имеют важную для формирования жизни на Земле особенность – при наличии в растворе одной полинуклеотидной цепочки спонтанно происходит процесс образования второй (параллельной) цепочки на основании комплементарного соединения родственных нуклеотидов. Одинаковое число нуклеотидов, в обоих цепочках и их химическое родство, является непременным условием для осуществления такого рода реакций. Однако при синтезе белка, когда информация с иРНК реализуется в структуру белка никакой речи о соблюдении принципа комплементарности идти не может. Это связано с тем, что в иРНК, и в синтезированном белке различно не только число мономеров, но и, что особенно важно, отсутствует структурное сходство между ними (с одной стороны нуклеотиды, с другой аминокислоты). Понятно, что в этом случае возникает необходимость создания нового принципа точного перевода информации с полинуклеотида в структуру полипептида. В эволюции такой принцип был создан и в его основу был заложен генетический код.
Генетический код – это система записи наследственной информации в молекулах нуклеиновых кислот, основанная на определённом чередовании последовательностей нуклеотидов в ДНК или РНК, образующих кодоны, соответствующие аминокислотам в белке.
Генетический код имеет несколько свойств.
Вырожденность или избыточность.
Следует отметить, что некоторые авторы предлагают ещё и другие свойства кода, связанные с химическими особенностями входящих в код нуклеотидов или с частотой встречаемости отдельных аминокислот в белках организма и т.д. Однако эти свойство вытекают из вышеперечисленных, поэтому там мы их и рассмотрим.
а. Триплетность. Генетический код, как и многое сложно организованные система имеет наименьшую структурную и наименьшую функциональную единицу. Триплет – наименьшая структурная единица генетического кода. Состоит она из трёх нуклеотидов. Кодон – наименьшая функциональная единица генетического кода. Как правило, кодонами называют триплеты иРНК. В генетическом коде кодон выполняет несколько функций. Во-первых, главная его функция заключается в том, что он кодирует одну аминокислоту. Во-вторых, кодон может не кодировать аминокислоту, но, в этом случае, он выполняет другую функцию (см. далее). Как видно из определения, триплет – это понятие, которое характеризует элементарную структурную единицу генетического кода (три нуклеотидов). Кодон – характеризует элементарную смысловую единицу генома – три нуклеотида определяют присоединение к полипептидной цепочки одной аминокислоты.
Элементарную структурную единицу вначале расшифровали теоретически, а затем её существование подтвердили экспериментально. И действительно, 20 аминокислот невозможно закодировать одним или двумя нуклеотидом т.к. последних всего 4. Три нуклеотида из четырёх дают 4 3 = 64 варианта, что с избытком перекрывает число имеющихся у живых организмах аминокислот (см.табл. 1).
Представленные в таблице 64 сочетания нуклеотидов имеют две особенности. Во-первых, из 64 вариантов триплетов только 61 являются кодонами и кодируют какую либо аминокислоту, их называют смысловые кодоны. Три триплета не кодируют
Кодоны информационной РНК и соответствующие им аминокислоты
О с н о в а н и я к о д о н о в
Нонсенс
Нонсенс
Нонсенс
Механизм действия стоп-кодонов отличается от механизма действия смысловых кодонов. Это следует из того, что для всех кодоны, кодирующие аминокислоты, найдены соответствующие тРНК. Для нонсенс-кодонов тРНК не найдены. Следовательно, в процессе остановки синтеза белка тРНК не принимает участие.
Кодон АУГ (у бактерий иногда ГУГ) не только кодируют аминокислоту метионин и валин, но и является инициатором трансляции.
б. Вырожденность или избыточность.
61 из 64 триплетов кодируют 20 аминокислот. Такое трёхразовое превышение числа триплетов над количеством аминокислот позволяет предположить, что в переносе информации могут быть использованы два варианта кодирования. Во-первых, не все 64 кодона могут быть задействованы в кодировании 20 аминокислот, а только 20 и, во-вторых, аминокислоты могут кодироваться несколькими кодонами. Исследования показали, что природа использовала последний вариант.
Его предпочтение очевидно. Если бы из 64 варианта триплетов в кодировании аминокислот участвовало только 20, то 44 триплета (из 64) оставались бы не кодирующими, т.е. бессмысленными (нонсенс-кодонами). Ранее мы указывали, насколько опасно для жизнедеятельности клетки превращение кодирующего триплета в результате мутации в нонсенс-кодон - это существенно нарушает нормальную работу РНК-полимеразы, приводя в конечном итоге к развитию заболеваний. В настоящее время в нашем геноме три кодона являются бессмысленными, а теперь представьте, что было бы если число нонсенс-кодонов увеличится в примерно в 15 раз. Понятно, что в такой ситуации переход нормальных кодонов в нонсенс-кодоны будет неизмеримо выше.
Код, при котором одна аминокислота кодируется несколькими триплетами, называется вырожденным или избыточным. Почти каждой аминокислоте соответствует несколько кодонов. Так, аминокислота лейцин может кодироваться шестью триплетами — УУА, УУГ, ЦУУ, ЦУЦ, ЦУА, ЦУГ. Валин кодируется четырьмя триплетами, фенилаланин — двумя и только триптофан и метионинкодируются одним кодоном. Свойство, которое связано с записью одной и той же информации разными символами носит названиевырожденность.
Число кодонов, предназначенных для одной аминокислоты, хорошо коррелируется с частотой встречаемости аминокислоты в белках.
И это, скорее всего, не случайно. Чем больше частота встречаемости аминокислоты в белке, тем чаще представлен кодон этой аминокислоты в геноме, тем выше вероятность его повреждения мутагенными факторами. Поэтому понятно, что мутированный кодон имеет больше шансов кодировать туже аминокислоту при высокой его вырожденности. С этих позиций вырожденность генетического кода является механизмом защищающим геном человека от повреждений.
Необходимо отметить, что термин вырожденность используется в молекулярной генетики и в другом смысле. Так основная часть информации в кодоне приходится на первые два нуклеотида, основание в третьем положении кодона оказывается малосущественным. Этот феномен называют “вырожденностью третьего основания”. Последняя особенность сводит до минимума эффект мутаций. Например, известно, что основной функцией эритроцитов крови является перенос кислорода от легких к тканям и углекислого газа от тканей к легким. Осуществляет эту функцию дыхательный пигмент — гемоглобин, который заполняет всю цитоплазму эритроцита. Состоит он из белковой части – глобина, который кодируется соответствующим геном. Кроме белка в молекулу гемоглобина входит гем, содержащий железо. Мутации в глобиновых генах приводят к появлению различных вариантов гемоглобинов. Чаще всего мутации связаны с заменой одного нуклеотида на другой и появлением в гене нового кодона, который может кодировать новую аминокислоту в полипептидной цепи гемоглобина. В триплете, в результате мутации может быть заменён любой нуклеотид – первый, второй или третий. Известно несколько сотен мутаций, затрагивающих целостность генов глобина. Около400из них связаны с заменой единичных нуклеотидов в гене и соответствующей аминокислотной заменой в полипептиде. Из них только100замен приводят к нестабильности гемоглобина и различного рода заболеваниям от легких до очень тяжелых. 300 (примерно 64%) мутаций-замен не влияют на функцию гемоглобина и не приводят к патологии. Одной из причин этого является упомянутая выше “вырожденность третьего основания”, когда замена третьего нуклеотида в триплете кодирующем серин, лейцин, пролин, аргинин и некоторые другие аминокислоты приводит к появлению кодона-синонима, кодирующего ту же аминокислоту. Фенотипически такая мутация не проявится. В отличие от этого любая замена первого или второго нуклеотида в триплете в 100 % случаях приводит к появлению нового варианта гемоглобина. Но и в этом случае тяжёлых фенотипических нарушений может и не быть. Причиной этому является замена аминокислоты в гемоглобине на другую сходную с первой по физико-химическим свойствам. Например, если аминокислота, обладающая гидрофильными свойствами, заменена на другую аминокислоту, но с такими же свойствами.
Гемоглобин состоит из железопорфириновой группы гема (к ней и присоединяются молекулы кислорода и углекислоты) и белка — глобина. Гемоглобин взрослого человека (НвА) содержит две идентичные -цепи и две -цепи. Молекула -цепи содержит 141 аминокислотных остатков, -цепочка — 146, - и -цепи различаются по многим аминокислотным остаткам. Аминокислотная последовательность каждой глобиновой цепи кодируется своим собственным геном. Ген, кодирующий -цепь располагается в коротком плече 16 хромосомы, -ген — в коротком плече 11 хромосомы. Замена в гене, кодирующем -цепь гемоглобина первого или второго нуклеотида практически всегда приводит к появлению в белка новых аминокислот , нарушению функций гемоглобина и тяжёлым последствия для больного. Например, замена “Ц” в одном из триплетов ЦАУ (гистидин) на “У” — приведет к появлению нового триплета УАУ, кодирующего другую аминокислоту — тирозин Фенотипически это проявится в тяжёлом заболевании.. Аналогичная замена в 63 положении -цепи полипептида гистидина на тирозин приведет к дестабилизации гемоглобина. Развивается заболевание метгемоглобинемия. Замена, в результате мутации, глутаминовой кислоты на валин в 6-м положении -цепи является причиной тяжелейшего заболевания — серповидно-клеточной анемии. Не будем продолжать печальный список. Отметим только, что при замене первых двух нуклеотидов может появится аминокислота по физико-химическим свойствам похожая на прежнюю. Так, замена 2-го нуклеотида в одном из триплетов, кодирующего глутаминовую кислоту (ГАА) в -цепи на “У” приводит к появлению нового триплета (ГУА), кодирующего валин, а замена первого нуклеотида на “А” формирует триплет ААА, кодирующий аминокислоту лизин. Глутаминовая кислота и лизин сходны по физико-химическим свойствам — они обе гидрофильны. Валин — гидрофобная аминокислота. Поэтому, замена гидрофильной глютаминовой кислоты на гидрофобный валин, значительно меняет свойства гемоглобина, что, в конечном итоге, приводит к развитию серповидноклеточной анемии, замена же гидрофильной глютаминовой кислоты на гидрофильный лизин в меньшей степени меняет функцию гемоглобина — у больных возникает легкая форма малокровия. В результате замены третьего основания новый триплет может кодировать туже аминокислоты, что и прежней. Например, если в триплете ЦАУ урацил был заменён на цитозин и возник триплет ЦАЦ, то практически никаких фенотипических изменений у человека выявлено не будет. Это понятно, т.к. оба триплета кодируют одну и туже аминокислоту – гистидин.
В заключении уместно подчеркнуть, что вырожденность генетического кода и вырожденность третьего основания с общебиологических позиция являются защитными механизмами, которые заложены в эволюции в уникальной структуре ДНК и РНК.
в. Однозначность.
Каждый триплет (кроме бессмысленных) кодирует только одну аминокислоту. Таким образом, в направлении кодон – аминокислота генетический код однозначен, в направлении аминокислота – кодон – неоднозначен (вырожденный).

Кодон аминокислота

г. Полярность
Считывание информации с ДНК и с иРНК происходит только в одном направлении. Полярность имеет важное значение для определения структур высшего порядка (вторичной, третичной и т.д.). Ранее мы говорили о том, что структуры низшего порядка определяют структуры более высшего порядка. Третичная структура и структуры более высокого порядка у белков, формируются сразу же как только синтезированная цепочка РНК отходит от молекулы ДНК или цепочка полипептида отходит от рибосомы. В то время когда свободный конец РНК или полипептида приобретает третичную структуру, другой конец цепочки ещё продолжает синтезироваться на ДНК (если транскрибируется РНК) или рибосоме (если транскрибируется полипептид).
Поэтому однонаправленный процесс считывания информации (при синтезе РНК и белка) имеет существенное значение не только для определения последовательности нуклеотидов или аминокислот в синтезируемом веществе, но для жёсткой детерминации вторичной, третичной и т.д. структур.
д. Неперекрываемость.
Код может быть перекрывающимся и не перекрывающимся. У большинства организмов код не перекрывающийся. Перекрывающийся код найден у некоторых фагов.
Сущность не перекрывающего кода заключается в том, что нуклеотид одного кодона не может быть одновременно нуклеотидом другого кодона. Если бы код был перекрывающим, то последовательность из семи нуклеотидов (ГЦУГЦУГ) могла кодировать не две аминокислоты (аланин-аланин) (рис.33,А) как в случае с не перекрывающимся кодом, а три (если общим является один нуклеотид) (рис. 33, Б) или пять (если общими являются два нуклеотида) (см. рис. 33, В). В последних двух случаях мутация любого нуклеотида привела бы к нарушению в последовательности двух, трёх и т.д. аминокислот.
Однако установлено, что мутация одного нуклеотида всегда нарушает включение в полипептид одной аминокислоты. Это существенный довод в пользу того, что код является не перекрывающимся.
Поясним это на рисунке 34. Жирными линиями показаны триплеты кодирующие аминокислоты в случае не перекрывающегося и перекрывающегося кода. Эксперименты однозначно показали, что генетический код является не перекрывающимся. Не вдаваясь в детали эксперимента отметим, что если заменить в последовательности нуклеотидов (см. рис.34 ) третий нуклеотид У (отмечен звёздочкой) на какой-либо другой то:
1. При неперекрывающемся коде контролируемый этой последовательностью белок имел бы замену одной (первой ) аминокислоте (отмечена звёздочками).
2. При перекрывающемся коде в варианте А произошла бы замена в двух (первой и второй) аминокислотах (отмечены звёздочками). При варианте Б замена коснулась бы трёх аминокислот (отмечены звёздочками).
Однако многочисленные опыты показали, что при нарушении одного нуклеотида в ДНК, нарушения в белке всегда касаются только одной аминокислоты, что характерно для неперекрывающегося кода.
ГЦУГЦУГ ГЦУГЦУГ ГЦУГЦУГ










ГЦУ ГЦУ ГЦУ УГЦ ЦУГ ГЦУ ЦУГ УГЦ ГЦУ ЦУГ
*** *** *** *** *** ***
Аланин – Аланин Ала – Цис – Лей Ала – Лей – Лей – Ала – Лей




А Б В
Не перекрывающийся код Перекрывающийся код
Рис. 34. Схема, объясняющая наличие в геноме не перекрывающегося кода (объяснение в тексте).
Неперекрываемость генетического кода связана с ещё одним свойством – считывание информации начинается с определённой точки – сигнала инициации. Таким сигналом инициации в иРНК является кодон, кодирующий метионин АУГ.
Следует отметить, что у человека всё-таки имеется небольшое число генов, которые отступают от общего правила и перекрываются.
е. Компактность.
ж. Универсальность.
Код един для всех организмов живущих на Земле. Прямое доказательство универсальности генетического кода было получено при сравнении последовательностей ДНК с соответствующими белковыми последовательностями. Оказалось, что во всех бактериальных и эукариотических геномах используется одни и те же наборы кодовых значений. Есть и исключения, но их не много.
Первые исключения из универсальности генетического кода были обнаружены в митохондриях некоторых видов животных. Это касалось кодона терминатора УГА, который читался так же как кодон УГГ, кодирующий аминокислоту триптофан. Были найдены и другие более редкие отклонения от универсальности.
МЗ. Генетический код – это система записи наследственной информации в молекулах нуклеиновых кислот, основанная на определённом чередовании последовательностей нуклеотидов в ДНК или РНК, образующих кодоны,
соответствующие аминокислотам в белке.Генетический код имеет несколько свойств.
Методы: частично-поисковый, проблемного изложения, объяснительно-иллюстративный.
- формирование у учащихся целостной системы знаний о живой природе, ее системной организации и эволюции;
-умения давать аргументированную оценку новой информации по биологическим вопросам;
-воспитание гражданской ответственности, самостоятельности, инициативности
Образовательные: о биологических системах (клетка, организм, вид, экосистема); истории развития современных представлений о живой природе; выдающихся открытиях в биологической науке; роли биологической науки в формировании современной естественнонаучной картины мира; методах научного познания;
Развитие творческих способностей в процессе изучения выдающихся достижений биологии, вошедших в общечеловеческую культуру; сложных и противоречивых путей развития современных научных взглядов, идей, теорий, концепций, различных гипотез (о сущности и происхождении жизни, человека) в ходе работы с различными источниками информации;
Воспитание убежденности в возможности познания живой природы, необходимости бережного отношения к природной среде, собственному здоровью; уважения к мнению оппонента при обсуждении биологических проблем
Личностные результаты обучения биологии:
1. воспитание российской гражданской идентичности: патриотизма, любви и уважения к Отечеству, чувства гордости за свою Родину; осознание своей этнической принадлежности; усвоение гуманистических и традиционных ценностей многонационального российского общества; воспитание чувства ответственности и долга перед Родиной;
2. формирование ответственного отношения к учению, готовности и способности обучающихся к саморазвитию и самообразованию на основе мотивации к обучению и познанию, осознанному выбору и построению дальнейшей индивидуальной траектории образования на базе ориентировки в мире профессий и профессиональных предпочтений, с учётом устойчивых познавательных интересов;
Метапредметные результаты обучения биологии:
1. умение самостоятельно определять цели своего обучения, ставить и формулировать для себя новые задачи в учёбе и познавательной деятельности, развивать мотивы и интересы своей познавательной деятельности;
2. овладение составляющими исследовательской и проектной деятельности, включая умения видеть проблему, ставить вопросы, выдвигать гипотезы;
3. умение работать с разными источниками биологической информации: находить биологическую информацию в различных источниках (тексте учебника, научно популярной литературе, биологических словарях и справочниках), анализировать и
Познавательные: выделение существенных признаков биологических объектов и процессов; приведение доказательств (аргументация) родства человека с млекопитающими животными; взаимосвязи человека и окружающей среды; зависимости здоровья человека от состояния окружающей среды; необходимости защиты окружающей среды; овладение методами биологической науки: наблюдение и описание биологических объектов и процессов; постановка биологических экспериментов и объяснение их результатов.
Регулятивные: умение самостоятельно планировать пути достижения целей, в том числе альтернативные, осознанно выбирать наиболее эффективные способы решения учебных и познавательных задач; умение организовывать учебное сотрудничество и совместную деятельность с учителем и сверстниками; работать индивидуально и в группе: находить общее решение и разрешать конфликты на основе согласования позиций и учёта интересов; формирование и развитие компетентности в области использования информационно-коммуникационных технологий (далее ИКТ-компетенции).
Коммуникативные: формирование коммуникативной компетентности в общении и сотрудничестве со сверстниками, понимание особенностей гендерной социализации в подростковом возрасте, общественно полезной, учебно-исследовательской, творческой и других видов деятельности.
Технологии: Здоровьесбережения, проблемного, развивающего обучения, групповой деятельности
Приемы: анализ, синтез, умозаключение, перевод информации с одного вида в другой, обобщение.
Что такое генетический код: общие сведения
Продолжить формирование знаний об информационной роли в клетке нуклеиновых кислот, раскрыв особенности строения молекул РНК;
Охарактеризовать виды РНК: транспортной, информационной, строение этих молекул и функции в клетке.
Добиться "понимания и усвоения учащимися сущности генетического кода, его свойств: специфичности/ универсальности; углубить знания о молекулах РНК.
Основные положения
Важнейшим событием добиологической эволюции является возникновение генетического кода в виде последовательности кодонов РНК, а затем и ДНК, которая оказалась способной сохранять информацию о наиболее удачных комбинациях аминокислот в белковых молекулах.
Появление первых клеточных форм ознаменовало начало биологической эволюции, начальные этапы которой характеризовались появлением эукариотических организмов, полового процесса и возникновением первых многоклеточных организмов.
РНК — так же, как ДНК, представляет собой полимер, мономером которого нуклеотиды, только вместо тимина в РНК присутствует урацил, а вместе дезоксирибозы — рибоза.
Каждой аминокислоте в полипептид почке соответствует комбинация из трёх нуклеотидов — триплет.
Привлеките внимание учащихся к особенностям строения молекул РНК вирусов, подчеркните их двуцепочечность в отличие от одноцепочечности эукариотических клеток
1.Особенности строения рибонуклеиновых кислот
2.Локализация нуклеиновых кислот в клетке
3.Участие РНК в реализации генетической информации
В любой клетке и организме все особенности анатомического, морфологического и функционального характера определяются структурой белков, которые входят в них. Наследственным свойством организма является способность к синтезу определенных белков. В молекуле ДНК аминокислоты расположены в полипептидной цепочке, от которой зависят биологические признаки. Для каждой клетки характерна своя последовательность нуклеотидов в полинуклеотидной цепи ДНК. Это и есть генетический код ДНК. Посредством его записывается информация о синтезе тех или иных белков.
Немного истории
Идея о том, что, возможно, генетический код существует, была сформулирована Дж.Гамовым и А.Дауном в середине двадцатого столетия. Они описали, что последовательность нуклеотидов, отвечающая за синтез определенной аминокислоты, содержит по меньшей мере три звена. Позже доказали точное количество из трех нуклеотидов (это единица генетического кода), которое назвали триплет или кодон. Всего нуклеотидов насчитывается шестьдесят четыре, потому что молекулы кислот, где происходит синтез белка или РНК, состоит из остатков четырех различных нуклеотидов. -

Что такое генетический код
Способ кодирования последовательности белков аминокислот благодаря последовательности нуклеотидов характерен для всех живых клеток и организмов. Вот что такое генетический код. В ДНК есть четыре нуклеотида: аденин - А; гуанин - Г; цитозин - Ц; тимин - Т. Они обозначаются заглавными буквами латинскими или (в русскоязычной литературе) русскими. В РНК также присутствуют четыре нуклеотида, однако один из них отличается от ДНК: аденин - А; гуанин - Г; цитозин - Ц; урацил - У. Все нуклеотиды выстраиваются в цепочки, причем в ДНК получается двойная спираль, а в РНК — одинарная. Белки строятся на двадцати аминокислотах, где они, расположенные в определенной последовательности, определяют его биологические свойства.
Свойства генетического кода.
В представленной таблице присутствуют не все имеющиеся аминокислоты. Гидроксипролин, гидроксилизин, фосфосерин, иодопроизводных тирозина, цистин и некоторые другие отсутствуют, так как они являются производными других аминокислот, кодирующихся м-РНК и образующихся после модификации белков в результате трансляции. Из свойств генетического кода известно, что один кодон способен кодировать одну аминокислоту. Исключением является выполняющий дополнительные функции и кодирующий валин и метионин, генетический код. ИРНК, находясь в начале с кодоном, присоединяет т-РНК, которая несет формилметион. По завершении синтеза он отщепляется сам и захватывает за собой формильный остаток, преобразуясь в остаток метионина. Так, вышеупомянутые кодоны являются инициаторами синтеза цепи полипептидов. Если же они находятся не в начале, то ничем не отличаются от других. -

Генетическая информация
Под этим понятием подразумевается программа свойств, которая передается от предков. Она заложена в наследственности как генетический код. Реализуется при синтезе белка генетический код РНК (рибонуклеиновыми кислотами): информационной и-РНК; транспортной т-РНК; рибосомальной р-РНК. Информация передается прямой связью (ДНК-РНК-белок) и обратной (среда-белок-ДНК). Организмы могут получать, сохранять, передавать ее и использовать при этом наиболее эффективно. Передаваясь по наследству, информация определяет развитие того или иного организма. Но из-за взаимодействия с окружающей средой реакция последнего искажается, благодаря чему и происходит эволюция и развитие. Таким образом в организм закладывается новая информация. -
Вычисление закономерностей молекулярной биологии и открытие генетического кода проиллюстрировали то, что необходимо соединить генетику с теорией Дарвина, на основе чего появилась синтетическая теория эволюции — неклассическая биология. Наследственность, изменчивость и естественный отбор Дарвина дополняются генетически определяемым отбором. Эволюция реализуется на генетическом уровне путем случайных мутаций и наследованием самых ценных признаков, которые наиболее адаптированы к окружающей среде.
Расшифровка кода у человека
В девяностых годах был начат проект Human Genome, в результате чего в двухтысячных были открыты фрагменты генома, содержащие 99,99% генов человека. Неизвестными остались фрагменты, которые не участвуют в синтезе белков и не кодируются. Их роль пока остается неизвестной.
Последняя открытая в 2006 году хромосома 1 является самой длинной в геноме. Более трехсот пятидесяти заболеваний, в том числе рак, появляются в результате нарушений и мутаций в ней. Роль подобных исследований трудно переоценить. Когда открыли, что такое генетический код, стало известно, по каким закономерностям происходит развитие, как формируется морфологическое строение, психика, предрасположенность к тем или иным заболеваниям, обмен веществ и пороки индивидов.
Вопросы для обсуждения
Что является наследственным материалом у некоторых вирусов, не содержащих ДНК? Как происходит реализация наследственной информации у них?
Каковы свойства генетического кода?
Каковы пути передачи наследственной информации в биологических системах?
Какова сущность процесса передачи наследственной информации из поколения в поколение и из ядра в цитоплазму, к месту синтеза белка?
Генетический код. Транскрипция
Гены, ДНК и хромосомы
Что такое гены?
ЕГЭ по биологии. Генетический код
Книга для учителя Сивоглазов В.И., Сухова Т.С. Козлова Т. А. Биология: общие закономерности .
Универсальный генетический код всех живых организмов почти не менялся на протяжении более, чем 3,5 млрд. лет после момента его окончательного формирования. Значит, его структура несёт след его возникновения. В связи с этим представляет интерес анализ различных особенностей генетического кода, которые могут дать ключ к пониманию его ранней эволюции. Данная статья является вступительной для серии автопубликаций, посвященных возникновению генетического кода, и с неё лучше всего начинать знакомство с этой темы.
Относится к разделу Молекулярная биология
1) Канонических аминокислот не 20, а только 19. Аминокислотой мы можем называть вещество, которое одновременно содержит аминогруппу -NH2 и карбоксильную группу - COOH. Дело в том, что мономер белка - пролин - аминокислотой не является, поскольку в нём вместо аминогруппы присутствует иминогруппа, поэтому пролин правильней называть иминокислотой. Однако в дальнейшем во всех статьях, посвящённых ГК, для удобства я буду писать о 20 аминокислотах, подразумевая указанный ньюанс. Структуры аминокислоты приведены на рис. 1.
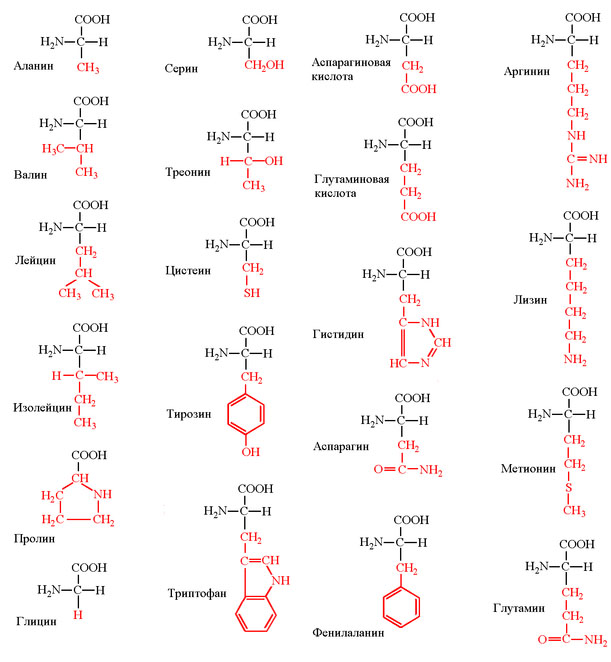
Рис. 1. Структуры канонических аминокислот. Аминокислоты имеют константные части, обозначенные на рисунке чёрным цветом, и вариабельные (или радикалы), обозначенные красным.
2) Соответствие аминокислот кодонам не всегда является однозначным. О нарушении случаев однозначности см. ниже.
Возникновение ГК означает возникновение кодируемого синтеза белка. Это событие является одним из ключевых для эволюционного формирования первых живых организмов.
Структура ГК представлена в круговой форме на рис. 2.
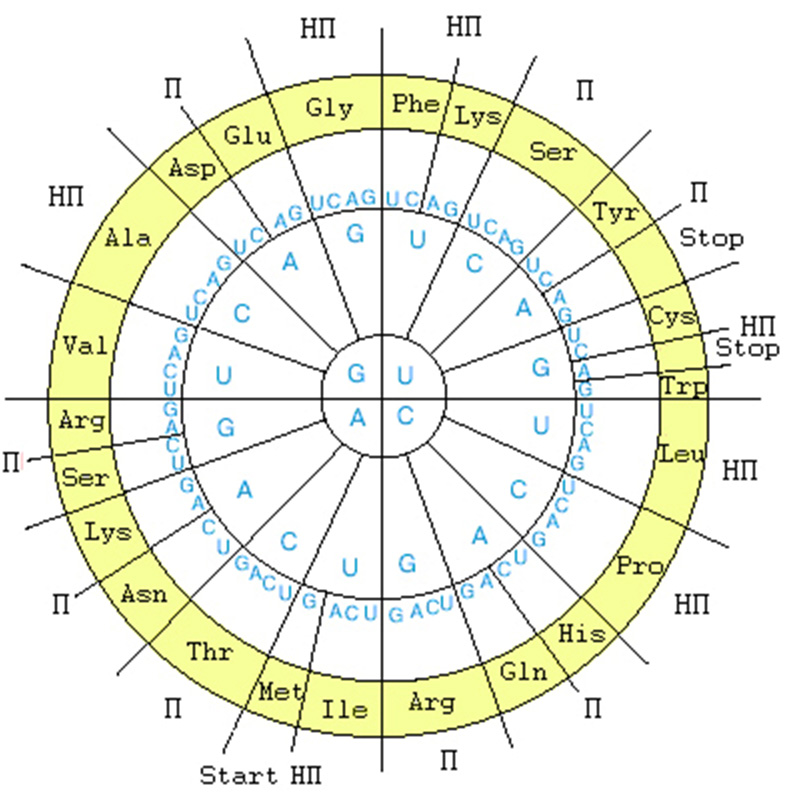
Рис. 2. Генетический код в круговой форме. Внутренний круг - первая буква кодона, второй круг - вторая буква кодона, третий круг - третья буква кодона, четвертый круг - обозначения аминокислот в трехбуквенном сокращении; П – полярные аминокислоты, НП – неполярные аминокислоты. Для наглядности симметрии важен избранный порядок символов U - C - A - G .
Итак, приступим к описанию основных свойств ГК.
1. Триплетность. Каждая аминокислота кодируется последовательностью из трёх нуклеотидов.
2. Наличие межгенных знаков препинания. К межгенным знакам препинания относятся последовательности нуклеиновой кислоты, на которых трансляци я начинается или заканчивается.
Трансляция может начаться не с любого кодона, а только со строго определённого – стартового. К стартовому кодону относится триплет AUG , с которого начинается трансляци я. В этом случае этот триплет кодирует или метионин, или другую аминокислоту – формилметионин (у прокариот), который может включаться только в начале синтеза белка. В конце каждого гена, кодирующего полипептид, находится, по меньшей мере, один из 3-х терминирующих кодонов, или стоп-сигналов: UAA, UAG, UGA. Они терминируют трансляци ю (так называется синтез белка на рибосоме).
3. Компактность, или отсутствие внутригенных знаков препинания. Внутри гена каждый нуклеотид входит в состав значащего кодона.
4. Неперекрываемость. Кодоны не перекрываются друг с другом, каждый имеет своё упорядочённое множестов нуклеотидов, которое не перекрывается с аналогичными множествами соседних кодонов.
5. Вырожденность. Обратное соответствие в направлении аминокислота-кодон неоднозначно. Это свойство называется вырожденностью. Серия – это множество кодонов, кодирующих одну аминокислоту, другими словами, это группа эквивалентных кодонов. Представим себе кодон в виде XYZ. Если XY определяет “ смысл ” (т.е. аминокислоту), то кодон называется сильным. Если же для определения смысл а кодона нужен определенный Z, то такой кодон называется слабым.
Вырожденность кода тесно связана с неоднозначностью спаривания кодон-антикодон (под антикодоном подразумевается последовательность из трёх нуклеотидов на тРНК , которая может комплементарно спариваться с кодоном на матричной РНК (см. более подробно об этом две статьи: Молекулярные механизмы обеспечения вырожденности кода и Правило Лагерквиста. Физико-химическое обоснование симметрий и соотношений Румера). Один антикодон на тРНК может узнавать отодного до трёх кодонов на мРНК.
6. Однозначность. Каждый триплет кодирует лишь одну аминокислоту или является терминатором трансляци и.
Известно три исключения.
Первое. У прокариот в первой позиции (заглавная буква) он кодирует формилметионин, а в любой другой - метионин. В начале гена формилметионин кодируется как обычным метиониновым кодоном AUG , так и ещё валиновым кодоном GUG или лейциновым UUG , которые внутри гена кодируют валин и лейцин, соответственно.
Во многих белках формилметионин отщепляется, либо удаляется формильная группа, в результате чего формилметионин превращается в обычный метионин.
Второе. В 1986 году сразу несколько групп исследователей обнаружили, что на мРНК терминирующий кодон UGA может кодировать селеноцистеин (см. рис. 3) при условии, что за ним следует особая последовательность нуклеотидов.
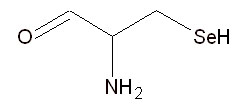
Рис. 3. Структура 21-й аминокислоты - селеноцистеина.
У E. coli (это латинское название кишечной палочки) селеноцистеил- тРНК в процессе трансляци и распознает в мРНК кодон UGA, но лишь в определенном контекст е: для узнавания UGA-кодона как о смысл енного важна последовательность длиной в 45 нуклеотидов, расположенная вслед за UGA-кодоном.
Рассмотренный пример показывает, что при необходимости живой организм может изменять смысл стандартного генетического кода. В этом случае генетическая информация, заключенная в генах, кодируется более сложным образом. Смысл кодона определяется в контекст е с определенной протяженной последовательностью нуклеотидов и при участии нескольких высокоспецифических белковых факторов. Важно, что селеноцистеиновая тРНК обнаружена в представителях всех трёх ветвей жизни (архей, эубактерий и эукариот), что указывает на древность происхождения селеноцистеинового синтеза, и возможно на присутствие его ещё в последнем универсальном общем предке (о нём речь пойдёт в других статьях). Скорей всего селеноцистеин встречается у всех без исключения живых организмов. Но в каждом отдельном организме селеноцистеин встречается не более, чем в паред есятков белков. Он входит в состав активных центров ферментов, в ряде гомологов которых на аналогичной позиции может функционировать обычный цистеин.
До недавнего времени считалось, что кодон UGA может считываться либо как селеноцистеин, либо кактерминальный, но недавно было показано, что у инфузории Euplotes кодон UGA кодирует или цистеин, илиселеноцистеин. См. " Генетический код допускает разночтения "
Третье исключение. У некоторых прокариот (5 видов архей и одной эубактерии - в Википедии информация сильно устарела) встречается особая кислота – пирролизин (рис. 4). Она кодируется триплетом UAG , который в каноническом коде служит терминатором трансляци и. Предполагается, что в этом случае, подобно случаю с кодированием селеноцистеина, считывание UAG как пирролизинового кодона происходит благодаря особой структуре на мРНК. Пирролизиновая тРНК содержит антикодон CTA и аминоацилируется АРСаз ой 2-го класса (про классификацию АРСаз см. статью "Кодазы помогают понять, как возник генетический код ").
UAG в качестве стоп-кодона используется редко, а если и используется, то часто за ним следует другой стоп-кодон.
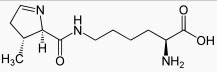
Рис. 4. Структура 22-й аминокислоты пирролизина.
7. Универсальность. После того, как в середине 60-х годов прошлого века расшифровка ГК была завершена, долгое время считалось, что код одинаков во всех организмах, что указывает на единство происхождения всего живого на Земле.
Попробуем понять, почему ГК универсален. Дело в том, что если бы в организме изменилось хотя бы одно правило кодирования, то это привело бы к тому, что изменилась структура значительной части белков. Такое изменение было бы слишком кардинальным и поэтому практически всегда летальное, так как изменение смысл а только одного кодона может затронуть в среднем 1/64 часть всех аминокислотных последовательностей.
Отсюда следует одна очень важная мысль - ГК почти не менялся со времени своего формирования более 3,5 млрд. лет назад. А, значит, его структура несёт в себе след его возникновения , и анализ этой структуры может помочь понять, как именно мог возникнуть ГК.
В действительности ГК может несколько отличаться у бактерий, митохондрий, ядерный код некоторых инфузорий и дрожжей. Cейчас насчитывают не менее 17 генетических кодов, отличающихся от канонического на 1-5 кодонов Суммарно во всех известных вариантах отклонений от универсального ГК используются 18 различных замен смысл а кодона. Больше всего отклонений от стандартного кода известно у митохондрий - 10. Примечательно, что митохондрии позвоночных, плоских червей, иглокожих, кодируются разными кодами, а плесневых грибков, простейших и кишечнополостных - одним.
Эволюционная близость видов - отнюдь не гарант того, чтобы у них были сходные ГК. Генетические коды могут различаться даже у разных видов микоплазм (одни виды имеют канонический код, а другие - отличающиеся). Аналогичная ситуация наблюдается и для дрожжей.
Важно отметить, что митохондрии - потомки симбиотических организмов, которые приспособились жить внутри клеток. Они имеют сильно редуцированный геном , часть генов переселилась в ядро клетки. Поэтому изменения ГК в них становятся уже не столь кардинальными.
Обнаруженные позднее исключения представляют особый интерес с точки зрения эволюции, поскольку могу помочь пролить свет на механизмы эволюции кода.
Митохондриальные коды у различных организмов.
Кодон
Универсальный код
Митохондриальные коды
Позвоночные
Беспозвоночные
Дрожжи
Растения
UGA
STOP
Trp
Trp
Trp
STOP
AUA
Ile
Met
Met
Met
Ile
CUA
Leu
Leu
Leu
Thr
Leu
AGA
Arg
STOP
Ser
Arg
Arg
AGG
Arg
STOP
Ser
Arg
Arg
Три механизма смены аминокислоты, кодируемой кодом.
Первый – когда какой-то кодон не используется (или почти не используется) каким-то организмом в силу неравномерности встречаемости каких-то нуклеотидов ( GC -состав), или комбинаций нуклеотидов. В результате такой кодон может вовсе исчезнуть из употребления (например, благодаря потере соответствующей тРНК ), а в дальнейшем может использоваться для кодирования другой аминокислоты без нанесения существенного ущерба организму. Этот механизм возможно отвечает за появление некоторых диалектов кодов у митохондрий.
Второй – превращение стоп-кодона в смысл овой. В этом случае часть у части транслируемых белков могут появиться дополнения. Однако ситуацию частично спасает то, что многие гены часто заканчиваются не одним, а двумя стоп-кодонами, поскольку возможны ошибки трансляци и, при которых стоп-кодоны считываются как аминокислоты.
Третий – возможно неоднозначное считывание определённых кодонов, как это имееют место у некоторых грибов.
8 . Связность. Группы эквивалентных кодонов (то есть кодонов, кодирующих одну и ту же аминокислоту) называются сериями. ГК содержит 21 серию, включая стоп-кодоны. В дальнейшем для определенности любая группа кодонов будет называться связной, если от каждого кодона этой группы можно перейти ко всем другим кодонам этой же группы путем последовательных замен нуклеотидов. Из 21 серии связны 18. 2 серии содержат по одному кодону, и лишь 1 серия для аминокислоты серин является несвязной и распадается на 2 две связные подсерии.
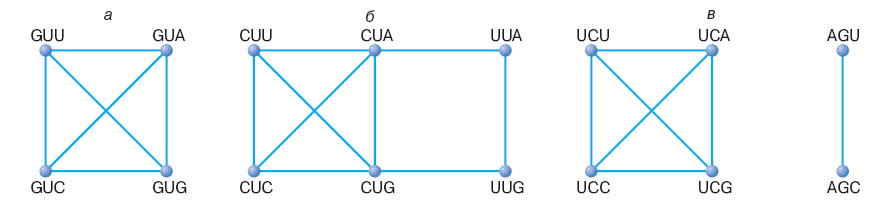
Рис. 5. Графы связности для некоторых кодовых серий. а - связная серия валина; б - связная серия лейцина; серия серина несвязная, распадается на две связных подсерии. Рисунок взят из статьи В.А. Ратнера " Генетический код как система ".
Свойство связности можно объяснить тем, что в период формирования ГК захватывал новые кодоны, которые минимально отличались от уже используемых.
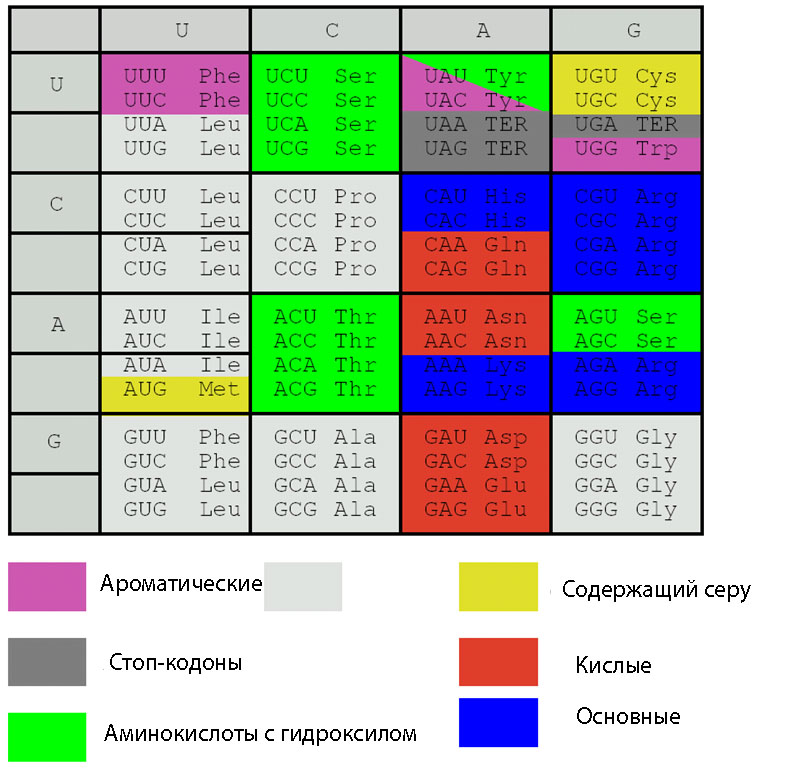
Рис. 6. Блочная структура ГК. Белым цветом обозначены аминокислоты с алкильной группой.
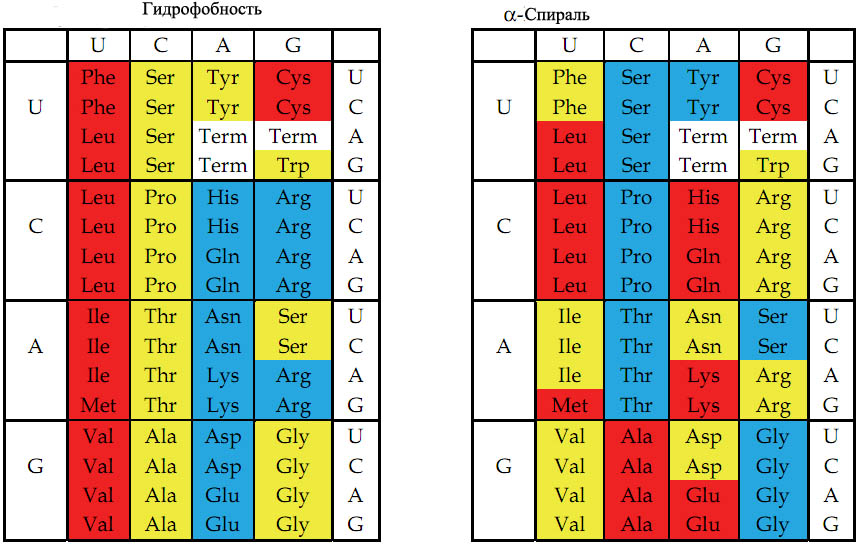
Рис. 7. Цветовое представление физико-химических свойств аминокислот, основанное на значениях, описанных в кн книге Стайерса "Биохимия" [Berg et al. 2002]. Слева - гидрофобность. Справа - способность к формированию альфа-спирали в белке. Красный, жёлтый и голубой цвета обозначают аминокислоты с большой, средней и малой гидрофобностью (слева) или соответствующей степенью способности к формированию альфа-спирали (справа).
Свойство блочности и регулярности также можно объяснить тем, что в период формирования ГК захватывал новые кодоны, которые минимально отличались от уже используемых.
Кодоны с одинаковыми первыми основаниями (приставками кодонов) кодируют аминокислоты с близкими путями биосинтеза [ Tailor F . J ., Coates D . 1989]. Кодоны аминокислот, принадлежащих к шикиматному , пируватному , аспартатному и глутаматному семействам, имеют в качестве приставок U, G, A и C, соответственно. О путях древнего биосинтеза аминокислот и его связи со свойствами современного кода см. "Древний дублетный генетический код был предопределён путями синтеза аминокислот". На основе этих данных некоторые исследователи делают вывод о том, что на формирование кода большое влияние оказали биосинтетические взаимоотношения между аминокислотами [ Wong J . T . 1975, Amirnovin R . 1997]. Однако сходство биосинтетических путей вовсе не означает сходство физико-химических свойств [ Di Giulio M . 1997 b ].
11. Помехоустойчивость. В самом общем виде помехоустойчивость ГК означает, что при случайных точковых мутациях и ошибках трансляци и не очень сильно меняются физико-химические свойства аминокислот.
Замена одного нуклеотида в триплете в большинстве случаев или не приводит к замене кодируемой аминокислоты, или приводит к замене на аминокислоту с той же полярностью.
Один из механизмов, обеспечивающих помехоустойчивость ГК – его вырожденность. Средняя вырожденность равна - число кодируемых сигналов/общее число кодонов, где к кодируемым сигналам относятся 20 аминокислот и знак терминации трансляци и. Усредненная вырожденность для всех аминокислот и знака терминации составляет три кодона на кодируемый сигнал.
Для того, чтобы количественно оценить помехоустойчивость, введём два понятия. Мутации замен нуклеотидов, не приводящие к смене класса кодируемой аминокислоты, называют консервативными. Мутации замен нуклеотидов, приводящие к смене класса кодируемой аминокислоты, называют радикальными .
Каждый триплет допускает 9 однократных замен. Всего кодирующих аминокислоты триплетов 61. Поэтому количество возможных замен нуклеотидов для всех кодонов –
61 x 9 = 549. Из них:
23 замены нуклеотидов приводят к появлению стоп-кодонов.
134 замены не меняют кодируемую аминокислоту.
230 замен не меняют класс кодируемой аминокислоты.
162 замены приводят к смене класса аминокислоты, т.е. являются радикальными.
Из 183 замен 3-его нуклеотида, 7 приводят к появлению терминаторов трансляци и, а 176 - консервативны.
Из 183 замен 1-ого нуклеотида, 9 приводят к появлению терминаторов, 114 - консервативны и 60 - радикальны.
Из 183 замен 2-го нуклеотида, 7 приводят к появлению терминаторов, 74 -консервативны, 102 - радикальны.
На основе этих расчётов получим количественную оценку помехоустойчивости кода, как отношение числа консервативных замен к числу радикальных замен. Оно равно 364/162=2.25
При реальной оценке вклада вырожденности в помехоустойчивость необходимо учитывать частоту встречаемости аминокислот в белках, которая варьирует в разных видах.
В чем причина помехоустойчивости кода? Большинство исследователей считают, что это свойство является следствием селекции альтернативных ГК [ Freeland S . J ., Hurst L . D . 1998].
Стивен Фриленд и Лоренс Херст генерировали случайные такие коды и выясняли, что только один из ста альтернативных кодов обладает не меньшей помехоустойчивостью по сравнению с универсальным ГК.
Еще более интересный факт обнаружился, когда эти исследователи ввели дополнительное ограничение, с тем чтобы учесть реально существующие тенденции в характере мутирования ДНК и появлении ошибок при трансляци и. При таких условиях лучше канонического кода оказался ТОЛЬКО ОДИН КОД ИЗ МИЛЛИОНА ВОЗМОЖНЫХ .
Столь беспрецедентную жизнестойкость генетического кода проще всего объяснить тем, что он сформировался в результате естественного отбора. Возможно когда-то в биологическом мире существовало множество кодов, каждый со своей чувствительностью к ошибкам. Организм, лучше справлявшийся с ними, имел больше шансов выжить, и канонический код просто победил в борьбе за существование. Это предположение кажется вполне реальным - ведь мы знаем, что альтернативные коды действительно существуют. Подробнее о помехоустойчивости см. Закодированная эволюция (С.Фриленд, Л. Херст "Закодированная эволюция".//В мире науки. – 2004, №7).
В заключение, предлагаю посчитать число возможных генетических кодов, которые можно сгенерировать для 20 канонических аминокислот. Почему-то это число нигде мне не попадалось. Итак, нам необходимо, чтобы в генерируемых ГК были обязательно 20 аминокислот и стоп-сигнал, кодируемые ХОТЯ БЫ ОДНИМ КОДОНОМ.
Мысленно будем нумеровать кодоны в каком-то порядке. Рассуждать будем следующим образом. Если у нас имеется ровно 21 кодон, то тогда каждая аминокислота и стоп-сигнал будут занимать ровно по одному кодону. В этом случае возможных ГК будет 21!
Если будет 22 кодона, то появляется лишний кодон, который может иметь один из любых 21 смысл ов, причём этот кодон может располагаться на любом из 22 мест, тогда как остальные кодоны имеют ровно по одному разному смысл у, как и для случая 21 кодонов. Тогда получим число комбинаций 21!х(21х22).
Если кодонов будет 23, то рассуждая аналогично, получим, что 21 кодон имеют ровно по одному разных смысл ов (21! вариантов), а два кодона - по 21 разных смысл а (21 2 смысл ов при ФИКСИРОВАННОМ положении этих кодонов). Число различных положений для этих двух кодонов будет 23х22. Общее число вариантов ГК для 23 кодонов - 21!х21 2 х23х22
Если кодонов будет 24 - то число ГК будет равно 21!х21 3 х24х23х22.
Если кодонов будет 64, то число возможных ГК будет 21!х21 43 х64!/21! = 21 43 х64! ~ 9.1х10 145

''Самым трудным в проблеме кода было понять, что код существует. На это потребовалось целое столетие. Когда это поняли, то для того чтобы разобраться в деталях, хватило каких-нибудь десяти лет'' [1].
Проблема генетического кода - это ключевая проблема. В конце 50-х - начале 60-х годов она приковывала к себе внимание, возбуждала активность умов, побуждала веру в величие и мудрость загадок науки. В широком смысле генетический код - это способ записи генетической информации в последовательностях нуклеиновых кислот (ДНК или РНК) о структуре полипептидов (белков). В конкретном смысле генетический код - это соответствие между триплетными кодонами матричной РНК (м-РНК) и аминокислотами кодируемого белка, задаваемое кодовой таблицей.
Развитие проблемы генетического кода прошло несколько этапов. Предтечами этой проблемы можно считать многих выдающихся исследователей. В частности, Н.К. Кольцов (1927, 1935) предложил в общей форме идею молекулы-гена и матричный принцип ее дублирования. Э. Шрёдингер (1944) явно сформулировал необходимость кодирования генетической информации в структуре генов-молекул. П. Колдуэлл и С. Хиншельвуд (1950) предложили идею матричного синтеза белков на ДНК. А. Даунс (1952) сформулировал гипотезу о синтезе белков на РНК.
Научные представления о генетическом коде как о реальной проблеме эксперимента и теории были сформулированы Г.А. Гамовым сразу же после обоснования Дж. Уотсоном и Ф. Криком (1953) модели строения двойной спирали ДНК. Первый этап изучения проблемы (1953-1961) можно назвать гипотетическим. Из модели Уотсона-Крика (см. подробнее [1]) вытекало представление о линейной последовательности ДНК - тексте, построенном из четырех типов нуклеотидов (A, T, G и C - четыре символа алфавита). Но кодируемые белки тоже имеют линейную первичную структуру - текст, построенный из 20 типов канонических аминокислот (алфавит из 20 символов). Поэтому Г.А. Гамов (1954) сразу же сформулировал идею генетического кода в конкретном смысле - как соответствие двух текстов, записанных при помощи двух разных алфавитов. Кроме того, он предложил использовать технические средства криптографии (расшифровки неизвестных кодов) для решения центральной проблемы генетики.
Генетический код сразу же приобрел облик великой загадки природы, ребуса для остроумных. Многие сотни математиков, физиков, химиков, биологов, включая Г.А. Гамова, Ф. Крика и др., предложили гипотетические варианты генетического кода, которые представляют теперь лишь исторический интерес. Реальный код оказался совсем иным.
Научными результатами первого этапа можно считать [1]: 1) постановку проблемы генетического кода; 2) формирование понятий линейного текста, алфавита для нуклеиновых кислот и белков, генетической информации, записанной в этих текстах при помощи символов алфавита; 3) представление о матричной роли РНК в трансляции; 4) понятие о кодонах и доказательство их неперекрывания; 5) предположение о триплетности кодонов и коллинеарности гена и белка, доказанное лишь в дальнейшем, и т.д.
Второй этап (1961-1966) можно назвать экспериментальным, так как в этот период генетический код был расшифрован в прямом эксперименте 1. В 1961 году Ф. Крик с сотрудниками в блестящей работе показали, что: а) кодоны триплетны; б) между ними нет разделительных знаков (''запятых''); в) гены, кодирующие структуру белков (цистроны), имеют фиксированное начало, ориентированное направление и фиксированный конец; г) существует небольшое число некодирующих триплетов (''нонсенсов'', бессмысленных кодонов), а код в целом сильно вырожден. В 1964 году Ч. Янофски с сотрудниками и С. Бреннер с сотрудниками показали, что ген и кодируемый им белок взаимно коллинеарны, то есть имеется последовательное соответствие между кодонами гена и аминокислотами белка.
Прямая расшифровка генетического кода in vitro оказалась возможной благодаря технике белкового синтеза в бесклеточных системах 3, то есть в клеточных экстрактах, содержащих все необходимые компоненты аппарата трансляции (т-РНК, рибосомы, аминокислоты, ферменты, источник энергии и т.д.), кроме м-РНК. Вводя в такие системы естественные м-РНК или искусственные небольшие олигорибонуклеотиды, можно было изучать специфичность включения меченых аминокислот в строящиеся полипептиды. М. Ниренберг и Ф. Ледер подавали в бесклеточную систему трансляции E. coli различные олигорибонуклеотиды и показали, что индивидуальные фракции тририбонуклеотидов, ассоциированные с рибосомами, связывают определенные фракции т-РНК, заряженные определенными мечеными аминокислотами. С помощью такого метода генетический код был расшифрован полностью. Летом 1966 года на симпозиуме по количественной биологии в Колд-Спринг-Харборе (США) все полученные данные были сведены Ф. Криком воедино [2]. Расшифрованный генетический код E. coli, исследованный in vitro, полностью согласовывался также с другими независимыми данными, полученными in vivo и для других видов. Этот вывод подтверждается также результатами секвенирования последних лет, когда найдено, что тысячи генов и кодируемых ими белков действительно соответствуют друг другу по правилам генетического кода.
Из 64 возможных триплетов 61 является смысловым кодоном, то есть кодирует аминокислоты. Все кодоны триплетны, неразрывны и не перекрываются в тексте, а также не разделены межкодонными знаками (запятыми). Все кодоны однозначны, то есть каждый кодирует единственную аминокислоту. Иначе говоря, в направлении кодон - аминокислота генетический код однозначен.
Обратное соответствие в направлении аминокислота - кодон неоднозначно, и это свойство называется вырожденностью. Отдельные аминокислоты кодируются группами (сериями) кодонов-синонимов. 18 серий из 20 содержат от двух до шести кодонов, две серии (Met и Trp) не вырождены, содержат по одному кодону. Средняя вырожденность генетического кода приблизительно три кодона на серию.
Вырожденность называется систематической, если кодоны-синонимы различаются в третьей позиции либо пуринами (R = A или G), либо пиримидинами (Y = U или C), либо вообще любыми из четырех нуклеотидов (N = A, G, U или C). Этим принципам удовлетворяют 30 пар кодонов из 32, а также восемь тетрад из 16. Все эти пары связны, а тетрады полносвязны. Остальные варианты вырожденности называют несистематическими. Они относятся к большим сериям: Leu и Arg - связные серии, Ser - несвязная серия, Ile - три кодона, полносвязная серия.
Генетический код содержит также знаки пунктуации (начала и конца) трансляции. Кодоны AUG, GUG и UUG у прокариот помимо кодирования аминокислот кодируют также инициацию трансляции. Однако однозначность кодирования при этом не нарушается, так как инициирующие знаки локализованы в определенном окружении (контексте), способном образовывать самокомплементарные ''шпильки''. У эукариот инициируют триплеты AUG и более слабо - UUG, AUA и ACG. Три ''вакантных'' триплета у E. coli - UAA (ochre), UAG (amber) и UGA (opal) - не кодируют аминокислот, а выполняют роль терминальных знаков трансляции (стоп-кодонов, нонсенс-кодонов или терминальных нонсенсов). В норме ими заканчиваются все цистроны, то есть транслируемые гены, единицы трансляции. Мутационное возникновение нонсенсов внутри гена приводит к преждевременной терминации трансляции и обрыву белка. Нонсенсы тоже образуют связную серию.
Расшифровка генетического кода была одним из самых выдающихся научных открытий ХХ века.
Третий этап изучения проблемы генетического кода (после 1966 года) связан с углубленным исследованием молекулярных механизмов кодирования, системных свойств генетического кода: симметрии, регулярности, помехоустойчивости, универсальности, а также путей его возникновения и эволюции (см. [4]). Молекулярной системой, обеспечивающей соответствие кодонов м-РНК и аминокислот, является набор адапторных молекул транспортных РНК (т-РНК) и набор кодирующих ферментов аминоацил-т-РНК-синтетаз (АРС-аз). Каждая специфическая молекула т-РНК имеет антикодон, взаимодействующий с кодоном м-РНК, а также специфический сайт взаимодействия с определенной АРС-азой и неспецифический сайт связывания аминокислоты. Каждая АРС-аза опознает все изоакцепторные (переносящие одну аминокислоту) фракции т-РНК, одну определенную аминокислоту и соединяет их макроэргической (энергобогатой) связью. Поэтому соответствие антикодона т-РНК и аминокислоты определяется именно АРС-азой. Фракции т-РНК выполняют функции адапторов (специфических посредников) между кодонами м-РНК и аминокислотами.
Многие свойства генетического кода обеспечиваются свойствами молекул т-РНК и АРС-аз. Триплетный и неразрывный антикодон выделен в антикодонной петле т-РНК специальными модифицированными нуклеотидами. Этим обеспечиваются триплетность и неразрывность узнаваемых кодонов матрицы. Все антикодоны одинаково триплетны, поэтому, начиная от инициирующего знака, трансляция осуществляется триплетными шагами, то есть формируется определенная рамка (фаза) трансляции - одна из трех возможных. В этом случае межкодоные знаки (запятые) не нужны, а кодоны не перекрываются. Инициирующие кодоны у E. coli опознаются специальной фракцией т-РНКF-Met, переносящей модифицированную аминокислоту формил-метионин. Терминальные нонсенсы вообще не имеют своих фракций т-РНК, а опознаются специальными белковыми факторами терминации.
Однозначность кода в направлении кодон - аминокислота обеспечивается строгой специфичностью АРС-аз. Каждая АРС-аза узнает единственную аминокислоту, поэтому неоднозначность исключена или маловероятна. В основе систематической вырожденности лежат правила неоднозначности спаривания кодон-антикодон, установленные Ф. Криком [1, 4]. Один антикодон может узнавать один, два или три кодона, различающиеся по третьей позиции. Согласно правилам неоднозначного спаривания, систематическая вырожденность в парах кодонов обеспечивается отдельными фракциями т-РНК, имеющими U, G или I (инозин) в трех позициях антикодонов. Вырожденность 3 у изолейцина (Ile) требует фракцию т-РНК с I в третьей позиции антикодона. Такой нуклеотид там действительно есть. Вырожденность 4 требует не менее двух фракций т-РНК, вырожденность 6 - не менее трех фракций. Всего генетический код E. coli требует не менее 32 фракций т-РНК. Реально у E. coli полное число генов т-РНК равно 86 для 79 фракций с различными антикодонами. Следовательно, многие фракции т-РНК частично дублируют друг друга.
Анализ таблицы генетического кода как целого позволяет выявить удивительные свойства его регулярности и симметрии [4]. Обозначим позиции нуклеотидов в кодонах (5'-1-2-3-3') через x - - y - z, где x - приставка, y - корень, z - окончание, а xy - основа кодона. Эти термины отражают сходные понятия лингвистики. Корни слов определяют их смысл. Все мутации, затрагивающие корень (y) кодона, также изменяют кодовую серию, то есть нарушают смысл кодона. Приставки слов тоже участвуют в определении смысла, хотя и не так жестко, а многие их изменения меняют смысл слов. Замены в приставках (x) кодонов чаще всего изменяют их смысл, но иногда являются синонимическими. Окончания слов обычно участвуют в словоизменении, то есть в синонимических преобразованиях. Аналогично 70% замен в окончаниях (z) кодонов синонимические. Наконец, приставка и корень слова образуют его основу, несущую полную или доминирующую смысловую нагрузку. Основа кодона (xy) тоже играет ключевую роль в особенностях генетического кода.
Регулярность генетического кода связана с распределением основных свойств кодонов и аминокислот по столбцам (корням) генетического кода. Кодоны характеризуются свойствами их основ и корней. Кодоны, имеющие одинаковые основы, образуют 16 тетрад генетического кода. Основа называется сильной, если она полностью определяет смысл (аминокислоту) кодонов тетрады. Нуклеотид в третьей позиции тоже, конечно, необходим, но он может быть любым (N). Таковы основы CU, GU, UC, CC, AC, GC, CG, GG. Легко заметить, что они содержат нуклеотиды в соотношении C : G : U : A = 7 : 5 : 3 : 1. Основа называется слабой, если для однозначного кодирования аминокислоты необходимо также участие определенного третьего нуклеотида (z) в кодоне. Таковы основы UU, AU, UA, CA, AA, GA, UG, AG. Они содержат нуклеотиды в обратном соотношении C : G : U : A = 1 : 3 : 5 : 7. Известно, что участки ДНК с избытком пар G-C более стабильны, чем A-T-богатые участки. Поэтому сильные основы образуют в среднем больше водородных связей с антикодонами т-РНК, чем слабые.
Аминокислоты имеют два основных характерных свойства, существенных в пространственной структуре глобулярных белков: размер (малые - М, средние - Ср, большие - Б) и полярность / неполярность (П / НП).
Легко заметить, что столбцы генетического кода, отвечающие определенным корням кодонов, имеют некоторые групповые свойства. Третий столбец (корень A) содержит полярные аминокислоты, немалые по размеру, а все основы кодонов слабые. Это как бы групповое свойство корня A. Второй столбец (корень C) содержит аминокислоты, малые по размеру, а все основы кодонов сильные. Первый столбец (корень U) включает неполярные аминокислоты разных (но не крайних) размеров, а свойства кодонов неоднозначны. Наконец, четвертый столбец (корень G) содержит все крайние и аномальные варианты аминокислот и кодовых серий [4]: самую реактивную аминокислоту (Cys), самую большую и плоскую (Trp), самую большую и корявую (Arg), самую маленькую (Gly), часть единственной несвязной серии (Ser) и неоднозначный терминальный нонсенс (UGA), который в ряде случаев кодирует 21-ю аминокислоту - селеноцистеин (Sec). Таким образом, если групповые свойства корней (и столбцов) отвечают каким-то общим правилам их возникновения, то четвертый столбец скорее напоминает свалку всего, что не попало в первые три столбца по групповым правилам их формирования.
Теперь рассмотрим не менее впечатляющее свойство симметрии генетического кода. Генетический код можно изобразить в круговой форме [4], где внутренний круг отвечает первым позициям кодонов, среднее кольцо - вторым позициям и внешнее кольцо - третьим позициям. Сильные основы изображены неподразделенными секторами внешнего кольца, а слабые - подразделенными. Свойство симметрии состоит в следующем:
1) проведем ось симметрии через центр круга перпендикулярно плоскости листа и повернем круг на 180° в плоскости листа. При этом все сильные и слабые основы сохраняют свои позиции, то есть совмещаются с одноименными;
2) проведем через центр плоскость симметрии, перпендикулярную плоскости листа и строкам текста. При зеркальном отражении круга в этой плоскости все сильные основы меняются местами со слабыми и наоборот;
3) проведем через центр плоскость симметрии, перпендикулярную плоскости листа и параллельную строкам текста. При зеркальном отражении круга в этой плоскости сильные основы меняются на слабые и наоборот.
Генетический код универсален в том смысле, что его основная часть одинакова для всех форм жизни на Земле. Этот вывод обоснован опытом массового секвенирования генов и белков. Почти всегда коллинеарное соответствие генов и белков согласуется с правилами генетического кода. Однако в некоторых экзотических системах трансляции (митохондрии животных, растений и грибов, хлоропласты растений, мельчайшие бактерии - микоплазмы, реснитчатые простейшие и др.) найдены минорные отклонения в генетическом коде, а также изменения правил неоднозначного спаривания и наборов антикодонов и фракций т-РНК. Это своеобразные ''диалекты'' генетического кода, отражающие специфику их эволюции и функционирования.
Несомненно, что генетический код явился продуктом добиологической молекулярной эволюции и продолжал частично эволюционировать в дальнейшем. В стохастическом процессе молекулярной эволюции свойства генетического кода могли быть: 1) либо предзаданы (предетерминированы) физико-химическими характеристиками компонент и условий, 2) либо отобраны как адаптивные среди альтернативных вариантов, 3) либо фиксированы случайно. Гипотезы возникновения генетического кода в разной степени учитывают эти возможности [3, 4].
Так, гипотеза ''замороженного случая'' (Ф. Крик, 1968 год) полагала, что исторически была фиксирована первая случайная, но удовлетворительная система кодирования, которая далее была размножена, подверглась эволюционному усложнению и оптимизации, так как обеспечивала ускоренное воспроизведение. Ясно, что крайний, чисто случайный вариант этой гипотезы нереален, поскольку код обладает очевидными неслучайными системными свойствами. Ясно также, что эти свойства отражают неслучайный, высоко организованный характер генетического кода, связанный с правилами синонимии кодовых серий.
Таким образом, генетический код E. coli представляет собой не случайный конгломерат соответствий между кодонами и аминокислотами, а высокоорганизованную систему соответствий, поддерживаемую сложными молекулярными механизмами. По выражению Френсиса Крика, внесшего решающий вклад в открытие и изучение кода, ''это ключ к молекулярной биологии, поскольку он показывает, как два великих языка полимеров - язык полинуклеотидов и язык полипептидов связаны между собой'' [2].
1. Ичас М. Биологический код. М.: Мир, 1971.
2. The Genetic Code. Cold Spring Harbor Symp. Quant. Biol. Cold Spring Harbor; N.Y. 1966. 31.
3. Молекулярная генетика. М.: Мир, 1963.
4. Ратнер В.А. Молекулярная генетика: Принципы и механизмы. Новосибирск: Наука, 1983.
Читайте также: