
Эксперименты показали что гены находящиеся в одной хромосоме наследуются сцепленно
Обновлено: 04.07.2024
Сцепление генов – это совместное наследование генов, расположенных в одной и той же хромосоме. Количество групп сцепления соответствует гаплоидному числу хромосом, то есть у дрозофилы 4. Природу сцепленного наследования объяснил Морган с сотрудниками. В качестве объекта исследования они избрали плодовую муху дрозофилу, которая оказалась очень удобной моделью для изучения данного феномена, так в клетках её тела находится только 4 пары хромосом и имеет место высокая скорость размножения (в течение года можно исследовать более 20-ти поколений). Итак, сцепленными признаками называются признаки, которые контролируются генами, расположенными в одной хромосоме. Естественно, что они передаются вместе в случаях полного сцепления (закон Моргана). Полное сцепление встречается редко, обычно – неполное, из-за влияния кроссинговера (перекрещивания и обмена участками гомологичных хромосом в процессе мейоза). То есть, гены одной хромосомы переходят в другую, гомологичную ей.
Частота кроссинговера зависит от расстояния между генами. Чем ближе друг к другу расположены гены в хромосоме, тем сильнее между ними сцепление и тем реже происходит их расхождение при кроссинговере, и, наоборот, чем дальше друг от друга отстоят гены, тем слабее сцепление между ними и тем чаще возможно его нарушение.

На рисунке 1 слева : расстояние между генами А и В маленькое, вероятность разрыва хроматиды именно между А и В невелика, поэтому сцепление полное, хромосомы в гаметах идентичны родительским (два типа), других вариантов не появляется.
На рисунке 1 справа: расстояние между генами А и В большое, повышается вероятность разрыва хроматиды и последующего воссоединения крест-накрест именно между А и В , поэтому сцепление неполное, хромосомы в гаметах образуются четырёх типов – 2 идентичные родительским (некроссоверные) + 2 кроссоверных варианта.
Количество разных типов гамет будет зависеть от частоты кроссинговера или расстояния между анализируемыми генами. Расстояние между генами исчисляется в морганидах – единицах расстояния между генами, находящимися в одной хромосоме. 1 морганида соответствует 1 % кроссинговера. Такая зависимость между расстояниями и частотой кроссинговера прослеживается только до 50 морганид. Частота кроссинговера между определенной парой генов – довольно постоянная величина (хотя радиация, химические вещества, гормоны, лекарства влияют на нее; например, высокая температура стимулирует кроссинговер).
Пример, основанный на опытах Моргана

Гены, расположенные в одной хромосоме, наследуются совместно. Фенотипы А – серое тело, нормальные крылья (повторяет материнскую форму). Б – тёмное тело, короткие крылья (повторяет отцовскую форму). В – серое тело, короткие крылья (отличается от родителей). Г – тёмное тело, нормальные крылья (отличается от родителей). В и Г получены в результате кроссинговера в мейозе.
Если скрестить мушку дрозофилу, имеющую серое тело и нормальные крылья (на рисунке самка), с мушкой, обладающей тёмной окраской и зачаточными (короткими) крыльями (на рисунке самец), то в первом поколении гибридов все мухи будут серыми с нормальными крыльями (А). Это гетерозиготы по двум парам аллельных генов, причём ген, определяющий серую окраску брюшка, доминирует над тёмной окраской, а ген, обусловливающий развитие нормальных крыльев, - доминирует над геном недоразвитых крыльев.
При анализирующем скрещивании гибрида F 1 с гомозиготной рецессивной дрозофилой (Б) подавляющее большинство потомков F 2 будет сходно с родительскими формами.
Это происходит потому, что гены, отвечающие за серое тело и нормальные крылья– сцепленные гены, также как и гены, отвечающие за тёмное тело и короткие крылья, т. е. они находятся в одной хромосоме. наследование сцепленных генов называют сцепленным наследованием.
Сцепление может нарушаться. Это доказывает наличие особей В и Г на рисунке, т. е. если бы сцепление не нарушалось, то этих особей бы не существовало, однако они есть. Это происходит в результате кроссинговера, который и нарушает сцепленность этих генов.

На рисунке 3 опыт Моргана отображен подробно. Несцепленное наследование: два гена находятся в разных хромосомах, гетерозигота с равной вероятностью дает четыре типа гамет:

Сцепленное наследование : два гена находятся в одной хромосоме.
а) При полном сцеплении гетерозигота дает только два типа гамет.
б) При неполном сцеплении гетрозигота дает четыре типа гамет, но не с равной вероятностью.

На вышесказанном строится хромосомная теория наследственности Моргана:
1. Гены находятся в хромосомах и расположены в линейной последовательности на определенных расстояниях друг от друга.
2. Гены, расположенные в одной хромосоме, составляют группу сцепления. Число групп сцепления равно гаплоидному числу хромосом. Признаки, гены которых находятся в одной хромосоме, наследуются сцепленно (т. е. в тех же сочетаниях, в которых они были в хромосомах исходных родительских форм).
3. Новые сочетания генов, расположенных в одной паре хромосом, могут возникать в результате кроссинговера в процессе мейоза. Частота кроссинговера зависит от расстояния между генами.
4. Учитывая линейное расположение генов в хромосоме и частоту кроссинговера как показателя расстояния между генами, можно построить карты хромосом. За единицу расстояния между генами принята частота кроссинговера равная 1 % (морганида, сантиморган, сМ).
Задачи на нахождение расстояния между генами
Решение задачи на определение вероятности рождения здоровый и больных детей
Наследование признаков, сцепленных с полом
Сцепленными с полом называются признаки, гены которых расположены не в аутосоме (неполовой хромосоме), а в гетеросоме (половой хромосоме). Схема решения задач на наследование признаков, сцепленных с полом, иная, чем на аутосомное моногибридное скрещивание. В случае, если ген сцеплен с Х-хромосомой, он может передаваться от отца только дочерям, а от матери в равной степени и дочерям, и сыновьям. Если ген сцеплен с Х-хромосомой и является рецессивным, то у самки он проявляется только в гомозиготном состоянии. У самцов второй Х-хромосомы нет, поэтому такой ген проявляется всегда.
При решении задач этого типа используются не символы генов (А, а, В, b), как при аутосомном наследовании, а символы половых хромосом X, Y с указанием локализованных в них генов (X А , X а ).
Аномалии, сцепленные с полом, чаще контролируются рецессивными генами, локализованы в Х-хромосоме и проявляются при генотипе ХY (т. е. у самцов млекопитающих и самок птиц).
Выше были рассмотрены примеры, где ген, сцепленный с полом, располагался в Х-хромосоме, но есть гены, локализованные в Y-хромосоме. У видов, у которых мужской пол гетерогаметен, этот ген может передаваться только самцам. У человека ген одного из видов синдактилии, выражающейся в образовании перепонки между 2 и 3 пальцами на ноге, локализован на Y-хромосоме, поэтому синдактилия возникает только у мужчин. Известна еще одна аномалия – гипертрихоз края ушной раковины (ряды волос на ухе), передающиеся по такому же механизму. В изучаемой семье с этой аномалией она передавалась в пяти поколениях по мужской линии.
Дальнейшие исследования генетиков показали, что законы Менделя о независимом наследовании признаков при дигибридном скрещивании применимы лишь тогда, когда разные гены располагаются в разных парах гомологичных хромосом. В том случае, если два гена находятся в одной паре гомологичных хромосом, расщепление в потомстве гибридов будет другим.
У любого организма генов значительно больше, чем хромосом. Например, у человека имеется около миллиона генов, а хромосом всего 23 пары. Следовательно, в одной хромосоме размещается в среднем несколько тысяч генов. Гены, расположенные в одной хромосоме, называют сцепленными. Все гены этой хромосомы образуют группу сцепления, которая при мейозе обычно попадает в одну гамету.
Значит, гены, входящие в одну группу сцепления, не подчиняются закону независимого наследования, а при дигибридном скрещивании вместо ожидаемого расщепления по фенотипу в соотношении 9:3:3:1 дают соотношение 3:1, как при моногибридном скрещивании.
Закономерности сцепленного наследования были установлены американским биологом Томасом Морганом (1866-1945). В качестве объекта он использовал плодовую муху дрозофилу. У дрозофилы окраску тела и длину крыльев определяют следующие пары аллелей: А - серое тело, а - черное тело, В - длинные крылья, b - зачаточные крылья. Гены, отвечающие за окраску тела и длину крыльев, находятся в одной паре гомологичных хромосом и наследуются сцепленно.

При скрещивании дрозофилы с серым телом и длинными крыльями с дрозофилой, имеющей черное тело и зачаточные крылья, все гибриды первого поколения имели серую окраску тела и длинные крылья.
При дальнейшем скрещивании между собой гибридных мух первого поколения в F2 не произошло ожидаемого расщепления по фенотипу 9:3:3:1. Вместо этого в F2 были получены мухи с родительскими фенотипами в соотношении примерно 3:1 . Появление в F2 двух фенотипов вместо четырех позволило сделать вывод, что гены окраски тела и длины крыльев дрозофил находятся в одной хромосоме. Так был установлен закон Т.Моргана: гены, расположенные в одной хромосоме, наследуются совместно - сцепленно, то есть наследуются преимущественно вместе.
Однако при дигибридном скрещивании при сцепленном наследовании признаков не всегда появляются особи только двух фенотипов. Иногда появляются особи еще двух фенотипов с перекомбинацией (новым сочетанием) родительских признаков: серое тело - зачаточные крылья, черное тело - длинные крылья. (Особей с такими фенотипами немного - около 8,5% каждого типа.) Почему же нарушается сцепление генов и появляются особи с новыми фенотипами? Было установлено, что сцепление генов может быть полным и неполным.
Полное сцепление наблюдается в том случае, если скрещиваются серый самец с длинными крыльями и самка с черным телом и зачаточными крыльями. Расщепление по фенотипу в этом случае будет 1:1, то есть наблюдается полное сцепление генов в одной хромосоме.
При скрещивании серой длиннокрылой самки с самцом, имеющим черное тело и зачаточные крылья, расщепление по фенотипу будет примерно 41,5:41,5:8,5:8,5, что характеризует неполное сцепление. Причина нарушения сцепления заключается в том, что в ходе мейоза происходит кроссинговер и гомологичные хромосомы обмениваются своими участками. В результате гены, расположенные в одной из гомологичных хромосом, оказываются в другой хромосоме. Возникают новые сочетания признаков.
У самцов дрозофил в мейозе кроссинговер не происходит, поэтому при скрещивании серого длиннокрылого самца дрозофилы с рецессивной самкой с черным телом и зачаточными крыльями сцепление будет полным. Неполное сцепление наблюдается в том случае, если самка гетерозиготна, а самец гомозиготен. В данном примере кроссинговер происходит примерно у 17% самок.
Таким образом, если не происходит перекреста хромосом и обмена генами, то наблюдается полное сцепление генов. При наличии кроссинговера сцепление генов бывает неполным. Благодаря перекресту хромосом возникают новые сочетания генов и признаков. Чем дальше друг от друга расположены гены в хромосоме, тем больше вероятность перекреста между ними и обмена участками хромосом.
Количество разных типов гамет будет зависеть от частоты кроссинговера или расстояния между анализируемыми генами. Расстояние между генами исчисляется в морганидах: единице расстояния между генами, находящимися в одной хромосоме, соответствует 1% кроссинговера. Такая зависимость между расстояниями и частотой кроссинговера прослеживается только до 50 морганид.
Сравните результаты скрещивание дрозофил:
а) Полное сцепление без кроссинговера
б) С частотой кроссинговера равной 17 %

Результатом исследований Т.Х.Моргана стало создание им хромосомной теории наследственности:
1. Гены располагаются в хромосомах; различные хромосомы содержат неодинаковое число генов, причем набор генов каждой из негомологичных хромосом уникален;
2. Каждый ген имеет определенное место (локус) в хромосоме; в идентичных локусах гомологичных хромосом находятся аллельные гены;
3. Гены расположены в хромосомах в определенной линейной последовательности;
4. Гены, локализованные в одной хромосоме, наследуются совместно, образуя группу сцепления; число групп сцепления равно гаплоидному набору хромосом и постоянно для каждого вида организмов;
5. Сцепление генов может нарушаться в процессе кроссинговера; это приводит к образованию рекомбинантных хромосом ;
6 . Частота кроссинговера является функцией расстояния между генами: чем больше расстояние, тем больше величина кроссинговера (прямая зависимость);
7. Каждый вид имеет характерный только для него набор хромосом - кариотип.
томас морган морганиды закон томаса моргана сцепленное наследование кроссинговер дрозофила дрозофилы хромосомная теория наследственности


Сцепленное наследование. Г. Мендель опубликовал результаты своих исследований в 1865 г., однако тогда его открытия остались незамеченными. Только в 1900 г. К-Корренс (Германия), Г. де Фриз (Голландия) и Э. Чер мак (Австрия) независимо друг от друга обнаружили у разных видов растений те же закономерности наследования признаков, что и Г. Мендель. Английский генетик У. Бэтсон подтвердил законы Менделя на животных. Переоткрытие законов Менделя вызвало глубокий интерес к изучению закономерностей наследования признаков и способствовало быстрому развитию генетики.
В 1902 г. немецкий цитолог и эмбриолог Т. Б о в е р и представил доказательства участия хромосом в процессах передачи наследственной информации. Он показал, например, что нормальное развитие морского ежа возможно лишь при наличии всех хромосом. Подобную связь заметил в 1903 г. и американский цитологУ. С эттон. Так получили обоснование предположения Менделя
о наследственных факторах, о наличии одинарного набора этих факторов в гаметах и двойного — в зиготах. В 1909 г. датский биолог В. Иогансен ввел понятие s.ген:/.
В 1910 г. американский генетик Т. Морган экспериментально доказал, что гены расположены в хромосомах. Многочисленные исследования Моргана и его учеников привели к целому ряду важнейших открытий, которые легли в основу хромосомной теории наследственности. Одно из ее положений можно сформулировать следующим образом: гены расположены в хромосомах в линейном порядке и занимают определенные участки — локусы, причем аллельные гены находятся в одинаковых локусах гомологичных хромосом.
Закон независимого наследования (третий закон Менделя) справедлив в том случае, если неаллельные гены находятся в разных парах хромосом. Однако количество генов у живых организмов значительно больше числа хромосом. Например, у человека около 25 тыс. генов, а количество хромосом —
23 пары (2п = 46); у плодовой мушки дрозофилы приблизительно 14 тыс. генов и всего 4 пары хромосом (2п = 8). Следовательно, каждая хромосома содержит множество генов. Будут ли гены, локализованные в одной хромосоме, наследоваться независимо? Очевидно, что нет.
Гены, расположенные в одной хромосоме, образуют группу сцепления и наследуются вместе. Совместное наследование генов Т. Морган предложил называть сцепленным наследованием (в отличие от независимого). Каждая пара гомологичных хромосом содержит гены, контролирующие одни и те же признаки, поэтому количество групп сцепления равно числу пар хромосом. Например, у человека 23 группы сцепления, а у дрозофилы — 4.

Вам известно, что при независимом наследовании дигетерозиготная особь, например =^=, образует четыре типа гамет в равном соотношении, т. е. по 25 %: Л В, АЬ, а В и ab. Это обусловлено тем, что неаллельные гены находятся в разных парах хромосом. Если же они расположены в гомологичных хромосомах, следовало бы ожидать, что дигетерозигота будет производить лишь два типа га-
мет: 50 % АВ и 50 % ab (обратите внимание на то, что сцепленные гены записываются в одну хромосому).
Однако Т. Морган обнаружил, что в большинстве случаев дигетерозиготные особи образуют не два, а четыре типа гамет. Помимо ожидаемых АВ и ab формируются также гаметы с новыми комбинациями генов: АЬ и аВ, только в меньшем процентном соотношении. Рассмотрим один из экспериментов Т. Моргана, в котором изучалось наследование сцепленных генов у дрозофилы.

Если почистить фрукты или овощи и не сразу выбросить очистки либо оставить фрукты на столе на несколько дней, то можно заметить, как вокруг остатков пищи начнут роиться маленькие мушки размером около 2—3,5 мм. Это дрозофилы — плодовые мушки, род насекомых отряда Двукрылые (рис. 95). Обычно дрозофилы имеют красные глаза и желтокоричневую окраску брюшка. Жизненный цикл дрозофил короток: развитие от яйца до половозрелой особи при 25 °С занимает 10 дней. Небольшие размеры, высокая плодовитость, простота культивирования и ряд других особенностей на долгое время сделали дрозофилу главным объектом генетики. Не один нобелевский лауреат, кроме своего интеллекта, обязан ей своими научными достижениями.
Путем скрещивания чистой линии дрозофил, имеющих серое тело и нормальные (длинные) крылья, с чистой линией, особи которой имели черное тело и зачаточные крылья, были получены гибриды первого поколения (рис. 96). Все они в соответствии с законом единообразия были серыми с нормально развитыми крыльями. Следовательно, у дрозофил серое тело (А) полностью доминирует над черным (а), а нормальные крылья (В) — над зачаточными (b). Все гибриды первого поколения — дигетерозиготы.
Затем было проведено анализирующее скрещивание (рис. 97). Дигетерозиготную самку из гибридного поколения скрестили с рецессивным дигомозиготным самцом (черное тело и зачаточные крылья). В потомстве было получено по 41,5 % особей с серым телом, нормальными крыльями и черным телом, зачаточными крыльями, а также по 8,5 % мух с серым телом, зачаточными крыльями и черным телом, нормальными крыльями.

Если бы гены, определяющие цвет тела и развитие крыльев, находились в разных парах хромосом, соотношение фенотипических классов было бы равным — по 25 %. Но этого не наблюдалось, значит, гены находятся в гомологичных хромосомах и наследуются сцепленно.

Несмотря на сцепление генов, АВ самка производила не два, а четыре типа гамет. Однако гамет с исходными сочетаниями сцепленных генов формировалось намного больше (АВ и ab вместе составили 83 %), чем с новыми их сочетаниями (сумма АЬ и дВ равна 17 %).
Было выяснено, что причиной появления хромосом с новыми комбинациями родительских генов является кроссинговер. Вы помните, что этот процесс происходит в профазе I мейоза и представляет собой обмен соответствующими участками между гомологичными хромосомами. Таким образом, кроссинговер препятствует полному (абсолютному) сцеплению генов. Гаметы, которые образуются в результате кроссинговера, и особи, которые развиваются при участии таких гамет, называются кроссоверными или рекомбинантными. В рассмотренном эксперименте гаметы АЬ и аВ являлись кроссоверными, а гаметы АВ и ab — некроссоверными (см. рис. 97).
Кроссинговер между конкретными сцепленными генами происходит с определенной вероятностью (частотой). Для расчета частоты кроссинговера (rf, от англ. recombination frequency — частота рекомбинации) можно пользоваться следующей формулой:
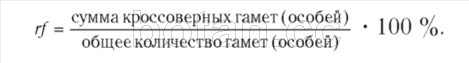
Таким образом, между генами А и В, контролирующими цвет тела и длину крыльев дрозофилы, кроссинговер происходит с частотой: rfAB= 17 %.
Дальнейшие исследования, проведенные Т. Морганом и его сотрудниками, показали, что частота кроссинговера пропорциональна расстоянию между генами, расположенными в одной хромосоме. Чем больше расстояние между сцепленными генами, тем чаще между ними происходит кроссинговер. И наоборот, чем ближе друг к другу расположены гены, тем меньше частота кроссинговера между ними. Чем объясняется эта закономерность?
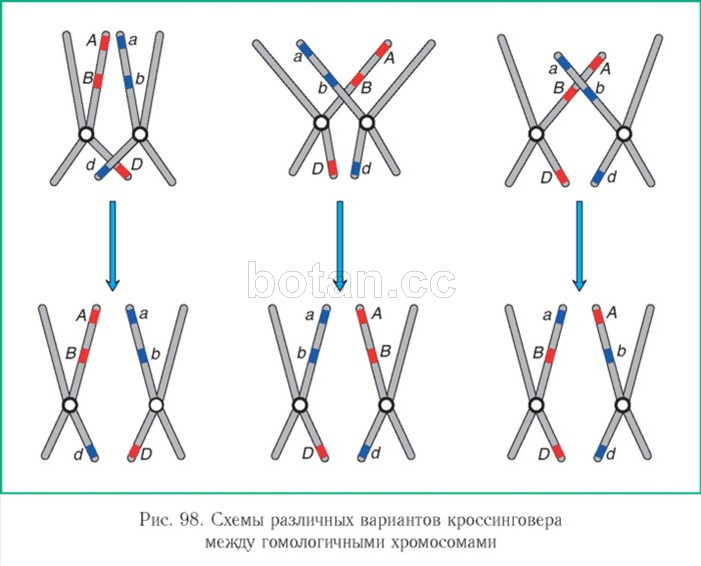
В профазе I мейоза при конъюгации гомологичных хромосом образование перекрестов между хроматидами осуществляется произвольно, на любых соответствующих участках. Рассмотрим рисунок 98.
Гены А и В (или а. и Ь) находятся сравнительно близко друг к другу. Вероятность того, что перекрест произойдет именно на участке, разделяющем эти гены, невелика. Гены А и D (или а. и d) располагаются на значительном расстоянии друг от друга. Поэтому вероятность того, что хроматиды перекрестятся на каком-либо участке между ними, намного выше. Значит, чем больше расстояние между генами, тем чаще они разделяются при кроссинговере.
Таким образом, частота кроссинговера позволяет судить о расстоянии между генами. В честь Т. Моргана единица измерения расстояния между генами получила название моргай и да или, что то же самое, санти моргай и да (сМ).
Морганида (сантиморганида, сМ) — это генетическое расстояние, на котором кроссинговер происходит с вероятностью 1 %.
Биологическое значение кроссинговера чрезвычайно велико. В результате этого процесса возникают новые комбинации родительских генов, что повышает генетическое разнообразие потомства и расширяет возможности адаптации организмов к различным условиям окружающей среды.
Генетические карты. Т. Морган и сотрудники его лаборатории показали, что знание частоты кроссинговера между сцепленными генами позволяет строить генетические карты хромосом. Генетическая карта представляет собой схему взаимного расположения генов, находящихся в одной группе сцепления, с учетом расстояний между ними (рис. 99).
Генетические карты хромосом уже составлены для человека, многих видов животных, растений, грибов и микроорганизмов. Наличие генетической карты свидетельствует о высокой степени изученности того или иного вида организма и представляет большой научный интерес. Такой организм является прекрасным объектом для проведения дальнейших экспериментальных работ, имеющих не только научное, но и практическое значение. В частности, знание генетических карт позволяет планировать работы по получению организмов с определенными сочетаниями признаков, что широко применяется в селекционной практике. Генетические карты хромосом человека используются в медицине для диагностики и лечения ряда наследственных заболеваний.

Основные положения хромосомной теории наследственности.
1. Гены в хромосомах расположены линейно, в определенной последовательности. Аллельные гены находятся в одинаковых локусах гомологичных хромосом.
2. Гены, расположенные в одной хромосоме, образуют группу сцепления и наследуются вместе. Количество групп сцепления равно числу пар хромосом.
3. Сцепление генов может нарушаться в результате кроссинговера, происходящего при конъюгации гомологичных хромосом в профазе I мейоза.
4. Частота кроссинговера пропорциональна расстоянию между генами: чем больше расстояние, тем выше частота кроссинговера, и наоборот.
Б. За единицу расстояния между сцепленными генами принята 1 морганида — расстояние, на котором кроссинговер происходит с вероятностью 1 %.
1. Всегда ли соблюдается III закон Г. Менделя? В каких случаях гены наследуются независимо, а в каких случаях — сцепленно?
2. Что такое группа сцепления? Чему равно количество групп сцепления в клетках разных организмов?
3. Почему частота кроссинговера между сцепленными генами зависит от расстояния между ними?
4. Что такое генетические карты хромосом? Каковы перспективы их использования?
5. Сформулируйте основные положения хромосомной теории наследственности.
6. Какие типы гамет и в каком процентном соотношении будут формировать дигетерозиготные особи - и ——, если известно, что расстояние между генами А и В составляет 20 морган ид?
7. Серый цвет тела у дрозофилы доминирует над желтым, красные глаза — над гранатовыми. Гены, отвечающие за эти признаки, локализованы в первой паре хромосом и находятся на расстоянии 44 морганиды. Скрестили чистые линии серотелых мух с гранатовыми глазами и желтотелых с красными глазами. Из полученных гибридов выбрали самку и подвергли ее анализирующему скрещиванию. Каким будет процентное соотношение фенотипических классов в потомстве?
8. У одного из видов растений рассеченные листья доминируют над цельными, а синяя окраска цветков — над розовой. В результате анализирующего скрещивания было получено потомство четырех фенотипических классов: 1) 133 растения с рассеченными листьями и синими цветками; 2) 362 растения с рассеченными листьями и розовыми цветками; 3) 127 растений с цельными листьями и розовыми цветками; 4) 378 растений с цельными листьями и синими цветками. Затем растения первого фенотипического класса скрестили с растениями второго фенотипического класса. Сколько процентов полученных гибридов будут иметь рассеченные листья и розовые цветки? Цельные листья и синие цветки?
Глава 1. Химические компоненты живых организмов
Глава 2. Клетка — структурная и функциональная единица живых организмов
Глава 3. Обмен веществ и преобразование энергии в организме
Глава 4. Структурная организация и регуляция функций в живых организмах
Глава 5. Размножение и индивидуальное развитие организмов
Глава 6. Наследственность и изменчивость организмов
Глава 7. Селекция и биотехнология
Биология: учеб. для 10-го кл. учреждений общ. сред, образования с рус. яз. обуч. / Н. Д. Лисов [и др.]; под ред. Н. Д. Лисова. — 3-е изд., перераб. — Минск : Народная асвета, 2014. — 270 с.: ил.

Из данного видеоурока вы узнаете о том, что в каждой хромосоме любого вида организма содержится большое количество генов, а также о том, что гены, расположенные в одной хромосоме, образуют группу сцепления. Вы выясните, что гены, входящие в группу сцепления, не подчиняются третьему закону Менделя о независимом наследовании. Узнаете о том, что сила сцепления между генами зависит от расстояния между ними. Познакомитесь с хромосомной теорией наследственности. Основные понятия урока: сцепленные гены, гетерозиготы, кроссоверные гаметы, рекомбинантные гаметы, хромосомная карта, хромосомная теория наследственности

В данный момент вы не можете посмотреть или раздать видеоурок ученикам
Чтобы получить доступ к этому и другим видеоурокам комплекта, вам нужно добавить его в личный кабинет, приобретя в каталоге.
Получите невероятные возможности



Конспект урока "Сцепленное наследование признаков. Хромосомная теория наследственности"
После того как Грегор Мендель открыл единые закономерности наследственности.
В начале 20-го века учёные генетики стали проводить множество экспериментов по скрещиванию на самых различных объектах. В итоге обнаружилось, что закономерности, установленные Менделем проявляются не всегда.
Мендель скрещивал дигетерозиготы – организмы которые отличались по двум признакам. Признаки, которые рассматривал Мендель были локализованы в разных гомологичных хромосомах.
Вспомним что третий закон Менделя формулируется так: каждая пара аллельных генов (и альтернативных признаков, контролируемых ими) наследуется независимо друг от друга.
Что значит независимо друг он друга?
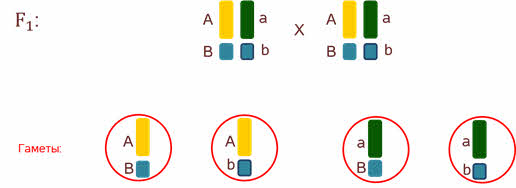
При скрещивании организмов 1-го поколения при мейозе образуются 4 типа гамет.
Где гены комбинируются в различных сочетаниях. Такие сочетания получились потому что гены находились в разных хромосомах.
Но в 1906 году Уильям Бэтсон и Риджинальд Пэннет, проводя скрещивание растений душистого горошка и анализируя наследование нескольких признаков формы пыльцы и окраски цветков, обнаружили, что эти признаки не дают независимого распределения в потомстве в соотношении 3:1, гибриды всегда повторяли признаки родительских форм.
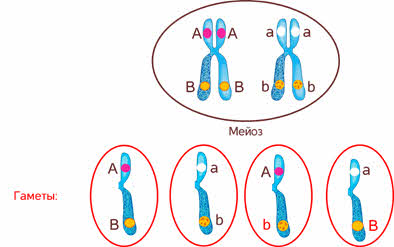
Стало ясно, что не для всех признаков характерно независимое распределение в потомстве и свободное комбинирование.
Дело в том, что ген формы пыльцы и ген окраски цветка лежат в одной хромосоме.
Признаков в организме значительно больше чем хромосом, в которых эти признаки локализованы. Следовательно, каждая хромосома несёт не один ген, а целую группу генов, отвечающих за развитие разных признаков.
Изучением наследования признаков, гены которых локализованы в одной хромосоме, занимался Томас Морган.
Он предложил закон сцепленного наследования (закон Моргана): гены, которые находиться в одной хромосоме, при мейозе попадают в одну гамету, то есть наследуются сцеплено.
Что значит сцеплено? То есть между генами, которые находиться в одной хромосоме возникают силы сцепления, то есть силы взаимодействия. И чем ближе эти гены, тем сильнее взаимодействие.
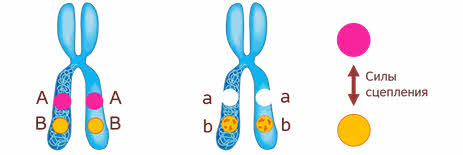
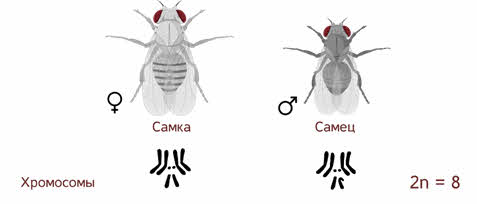
Если Мендель проводил свои опыты на горохе, то для Моргана основным объектом стала фруктовая мушка дрозофила, которая имела диплоидный набор из 8 хромосом.
Небольшие размеры, короткий жизненный цикл и простота культивирования позволяет использовать ряд видов дрозофил как образцовые объекты генетических исследований.
Самец и самка внешне хорошо различимы — у самца брюшко меньше и темнее.
Дрозофила фруктовая — наиболее важный для научных исследований вид дрозофил. Главными её характеристиками как модельного объекта является малое число хромосом. Дрозофила каждые две недели при температуре 25 °С достаточно легко размножаются в пробирках и даёт многочисленное потомство.
Рассмотрим один из первых экспериментов Томаса Моргана по изучению сцепленного наследования.
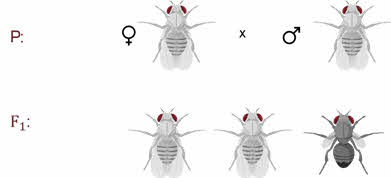
Скрещивая мушку дрозофилу с серым телом и нормальными крыльями с мушкой, имеющей темную окраску тела и зачаточные крылья, в первом поколении Морган получал гибриды, которые имели серое тело и нормальные крылья.
Ген А-большое отвечает за серое тело, рецессивный ген а-малое за чёрную окраску тела, доминантный ген В-большое за развитие длинных крыльев, а рецессивный ген бэ-малое за не развитие крыльев, то есть крылья остаются в зачаточном состоянии.
Значит ген, определяющий серую окраску брюшка, доминирует над темной окраской, а ген, обуславливающий развитие нормальных крыльев, — над геном недоразвитых.
Далее при скрещивании двух гетерозигот которые имеют серое тело и длинные крылья в первом поколении получается два фенотипических класса организмов.
Которые имеют серое тело и длинные крылья и чёрное тело с зачаточными крыльями.
То есть здесь расщепление идёт именно на 2 класса, а не на четыре как при дигибридном скрещивании Менделя.
Почему два? Дело в том, что гены окраски тела и длинны крыльев сцеплены в хромосоме.
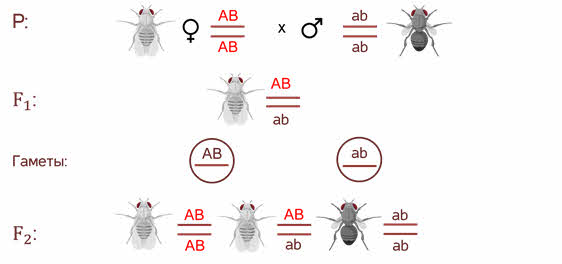
Символы АB АB ab аb располагаются не рядом как мы записывали их ранее, а как бы друг под другом с двумя чёрточками. Чёрточками мы условно обозначаем хромосомы.
В первом поколении организм гетерозиготен по обоим генам, но при образовании гамет эта гетерозиготность не даёт всех возможных комбинаций. То есть родительские гены остаются связанные между собой и гаметы получаются 2х типов.
При комбинации такого рода гамет в потомстве возникают всего 3 генотипических класса потомков.
Морган исследуя наследование сцепленных генов обнаружил что существует нарушение этого правила по дигибридному скрещиванию Менделя.
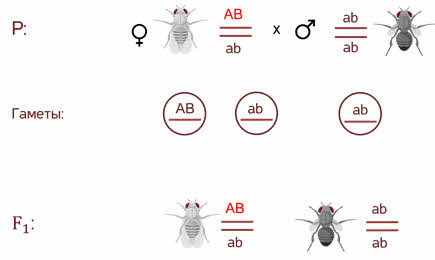
Он провёл анализирующее скрещивание.
Взял дигетерозиготную особь, которая получилась при скрещивании в первом поколении и скрестил её с чёрной мушкой с зачаточными крыльями, то есть оба рецессивных признака. У него получился необычный результат.
Морган рассуждал. Если гены окраски тела и формы крыльев локализованы в одной хромосоме, то при данном скрещивании должны были получиться две группы особей, повторяющие признаки родительских форм, так как материнский организм должен образовывать гаметы только двух типов — АB и аb, а отцовский — один тип — аb. Следовательно, в потомстве должны образовываться две группы особей, имеющих генотип АB аb и аa BB.
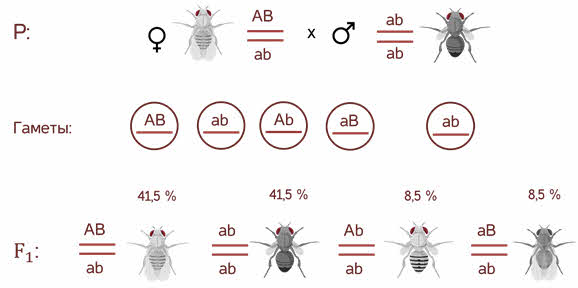
Однако в потомстве появляются особи (пусть и в незначительном количестве) с перекомбинированными признаками, то есть имеющие генотип Аa bb и aa Bb.
В потомстве явно преобладали особи с признаками родительских форм (41,5% были серые длиннокрылые и 41,5% — черные с зачаточными крыльями), и лишь незначительная часть мушек имела иное, чем у родителей, сочетание признаков (8,5% были серые с зачаточными крыльями и 8,5% — черные длиннокрылые).
Такие результаты могли быть получены только в том случае, если гены, отвечающие за окраску тела и форму крыльев, находятся в одной хромосоме.
В профазе первого мейотического деления гомологичные хромосомы (то есть идентичные хромосомы одной пары) конъюгируют (сближаются), и могут разрываться в месте контакта в этот момент между ними может произойти обмен участками – кроссинговер.
Кроссинго́вер или перекрёст — это процесс обмена участками гомологичных хромосом во время конъюгации в профазе первого мейоза. В результате образуются крассоверные гаметы.
Организмы, которые возникают в результате слияния кроссоверных гамет называются рекомбинантными.
Так в результате, кроссинговера в некоторых клетках происходит обмен участками хромосом между генами А и В, появляются гаметы Аb и аB, и, как следствие, в потомстве образуются четыре группы фенотипов, как при свободном комбинировании генов.
Однако кроссинговер происходит не после каждой конъюгации (сближения хромосом). И определить в каких участках хромосом он произойдет достаточно сложно.
В ходе эксперимента Томасу Моргану удалось доказать, что частота кроссинговера между генами прямо пропорциональна расстоянию между ними в хромосоме. То есть можно сказать что, чем дальше гены находятся друг от друга в хромосоме, тем чаще между ними происходит кроссинговер.
Если рассматривать 2 гена А и В мы можем увидеть 2 случая.
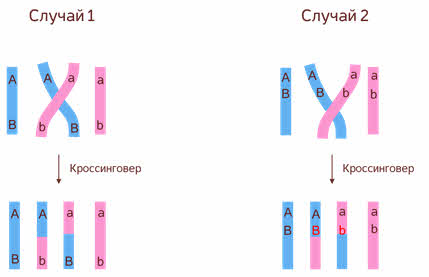
В первом случае гены А и В находиться по разные стороны перекрёста. Тогда после прохождения кроссинговера мы увидим новые сочетания аллелей этих двух генов. В данном случае Аb и аB.
Во 2м случае гены А и B находиться по одну сторону от перекрёста. Тогда после прохождения кроссинговера новых сочетаний аллелей данных двух генов мы не увидим.
Таким образом существуют понятия полного и неполного сцепления.
Неполное сцепление — это разновидность сцепленного наследования, при котором гены анализируемых признаков располагаются на некотором расстоянии друг от друга, что делает возможным кроссинговер между ними.
Полное сцепление — это разновидность сцепленного наследования, при котором гены анализируемых признаков располагаются так близко друг к другу, что кроссинговер между ними становится невозможным.
Это открытие позволило лаборатории Томаса Моргана разработать метод. Который позволяет построить хромосомные карты.
Хромосомные карты — это графическое изображение хромосомы, на котором определенные локусы (гены) отмечены соответственно расстоянию между ними.
Хромосомные карты составляют при помощи генетического анализа, который позволяет точно определить местоположение в хромосоме любого гена.
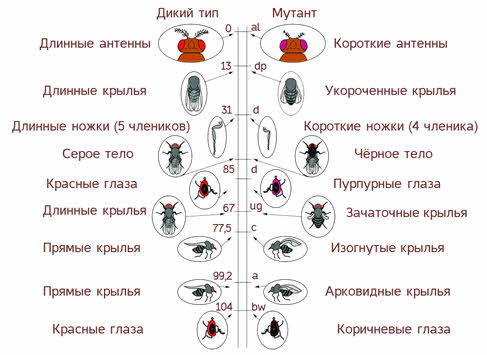
Хромосомная теория наследственности
Изучение Морганом наследования родительских признаков гибридами дрозофилы показало, что число групп сцепленного наследования было равным количеству пар гомологичных хромосом.
Например, у человека 46 хромосом, следовательно, 23 группы сцепления. У дрозофилы 8 хромосом, то есть 4 группы сцепления.
На этом основании был сделан вывод о строгой локализации конкретных генов в определенных парах хромосом.
Возникновение кроссоверных (рекомбинантных) особей дрозофилы можно было объяснить только линейным расположением генов в хромосомах и их обменом при кроссинговере в профазе первого мейоза.
Томас Морган обосновал хромосомную теорию наследственности. Согласно этой теории, передача наследственной информации связана с хромосомами, в которых линейно, в определенной последовательности, локализованы гены. Таким образом, именно хромосомы представляют собой материальную основу наследственности.
Формированию хромосомной теории способствовали данные, полученные при изучении генетики пола, когда были установлены различия в наборе хромосом у организмов различных полов.
Хромосомная теория наследственности сформулирована в 1911 г. американским учёным Томасом Морганом. Её сущность заключается в следующем:
· Основным материальным носителем наследственности являются хромосомы с локализованными в них генами.
· Гены в хромосомах расположены линейно, каждый ген имеет определенное место (локус) в хромосоме;
· Гены, расположенные в одной хромосоме, образуют группу сцепления и наследуются совместно;
· Число групп сцепления равно гаплоидному набору хромосом у гомогаметных особей и n+1 у гетерогаметных особей.
· Между гомологичными хромосомами может происходить обмен участками (кроссинговер); в результате кроссинговера возникают гаметы, хромосомы которых содержат новые комбинации генов.
· Сцепление генов может нарушаться в результате кроссинговера;
· Частота кроссинговера между гомологичными хромосомами зависит от расстояния между генами, локализованными в одной хромосоме. Чем это расстояние больше, тем выше частота кроссинговера.
Значение этой теории заключается в том, что она дала объяснение законам Менделя, вскрыла цитологические основы наследования признаков и генетические основы теории естественного отбора.
Читайте также: