
Что такое сцепленное наследование гдз
Обновлено: 04.07.2024
Явление сцепленного наследования и его цитологические основы
Закон независимого комбинирования генов основывается на тех положениях, что гены, определяющие те или иные черты и признаки, локализованы в гомологических хромосомах, а гены, кодирующие разные черты находятся в разных хромосомах. Но количество признаков намного превышает количество хромосом в живых организмах. Из этого следует логичный вывод, что каждый организм имеет число генов, которые способны независимо комбинироваться в мейозе, но ограничены числом пар хромосом. Вследствие этого на каждую хромосому приходится далеко не по одному гену.
Хромосомы наследуются как единое целое. Они сохраняют свою целостность при конъюгации и расхождении в мейозе. Поэтому гены, содержащиеся в одной хромосоме, как правило, наследуются совместно.
Гены, которые локализованы в одной хромосоме и способны наследоваться совместно, составляют группу сцепления. А совместное наследование генов соответственно называется сцеплением генов.
У организмов определенного вида количество групп сцепления равно количеству хромосом в гаплоидном наборе.
Хромосомная теория наследственности
Впервые явление сцепленного наследования признаков было описано в $1906$ году В. Бетсоном и Р. Пеннетом в опытах, проводимых с душистым горошком. Но они не смогли объяснить результаты опытов и пришли к выводу об ограниченности правила независимого комбинирования признаков, установленного Г. Менделем.
Экспериментальными исследованиями явления сцепленного наследования успешно занимался выдающийся американский естествоиспытатель и генетик Томас Хант Морган. Он со своими ассистентами и сотрудниками А. Стервантом, Г. Миллером и К. Бриджесом провел основательные исследования. Результаты этих исследований позволили предложить и аргументированно обосновать хромосомную теорию наследственности.
Готовые работы на аналогичную тему
Опыты Т. Х. Моргана
Для проведения исследований Т.Х.Морган избрал в качестве объекта муху-дрозофилу. С тех пор эта муха стала классическим объектом для различных генетических экспериментов. Их легко содержать, они быстро размножаются. А небольшое количество хромосом облегчает наблюдение.
Был проведен следующий опыт. Самцов дрозофилы, которые были гомозиготными по доминантным признакам окраски тела и формы крыльев (а именно - серое тело и нормальные крылья), ученые скрестили с самками, гомозиготными по рецессивным признакам (черное тело и недоразвитые крылья). Генотипы исследуемых особей обозначили соответственно ЕЕVV и ееvv. Всем гибридам первого поколения характерно было серое тело и нормальные крылья. Они были гетерозиготными. Их генотип можно было записать как EeVv. Затем провели анализирующее скрещивание. Для этого гибриды первого поколения скрестили с гомозиготами по рецессивным признакам. Теоретически можно было предположить, что произойдет расщепление признаков и пропорция полученных результатов будет выглядеть так: $1 : 1 : 1 : 1$. Другими словами каждого варианта будет примерно по $25$%. На самом же деле $41,5$% особей имели серое тело и нормальные крылья, $41,5$% - черное тело и недоразвитые крылья, $8,5$% - серое тело и недоразвитые крылья, $8,5$% - черное тело и нормальные крылья. Результаты опытов позволили Моргану сформулировать два важных предположения.
- Гены, которые определяют цвет тела и форму крыльев локализованы в одной хромосоме и в дальнейшем наследуются сцеплено.
- В процессе мейоза и образования гамет гомологические хромосомы некоторых особей обменялись участками и образовали новую группу сцепления.
Явление кроссинговера
Явление перекреста хромосом во время мейоза и последующий обмен участками хромосом получил название кроссинговера.
Он увеличивает комбинативную изменчивость, способствую появлению новых сочетаний аллелей. Были установлены следующие закономерности кроссинговера:
- Сила сцепления между двумя генами, которые расположены в одной хромосоме, обратно пропорциональна расстоянию между ними.
- Частота кроссинговера, который происходит между двумя сцепленными генами, это относительно постоянная величина для каждой конкретной пары генов.
Главным выводом моргановской гипотезы было то, что гены расположены в хромосоме по всей ее длине один за другим в линейном порядке.

Из данного видеоурока вы узнаете о том, что в каждой хромосоме любого вида организма содержится большое количество генов, а также о том, что гены, расположенные в одной хромосоме, образуют группу сцепления. Вы выясните, что гены, входящие в группу сцепления, не подчиняются третьему закону Менделя о независимом наследовании. Узнаете о том, что сила сцепления между генами зависит от расстояния между ними. Познакомитесь с хромосомной теорией наследственности. Основные понятия урока: сцепленные гены, гетерозиготы, кроссоверные гаметы, рекомбинантные гаметы, хромосомная карта, хромосомная теория наследственности

В данный момент вы не можете посмотреть или раздать видеоурок ученикам
Чтобы получить доступ к этому и другим видеоурокам комплекта, вам нужно добавить его в личный кабинет, приобретя в каталоге.
Получите невероятные возможности



Конспект урока "Сцепленное наследование признаков. Хромосомная теория наследственности"
После того как Грегор Мендель открыл единые закономерности наследственности.
В начале 20-го века учёные генетики стали проводить множество экспериментов по скрещиванию на самых различных объектах. В итоге обнаружилось, что закономерности, установленные Менделем проявляются не всегда.
Мендель скрещивал дигетерозиготы – организмы которые отличались по двум признакам. Признаки, которые рассматривал Мендель были локализованы в разных гомологичных хромосомах.
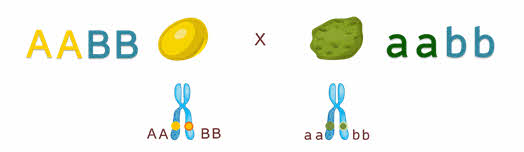
Вспомним что третий закон Менделя формулируется так: каждая пара аллельных генов (и альтернативных признаков, контролируемых ими) наследуется независимо друг от друга.
Что значит независимо друг он друга?
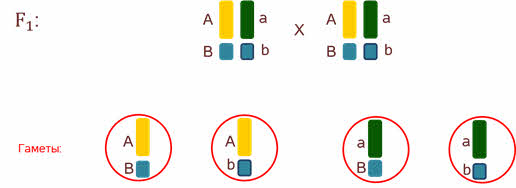
При скрещивании организмов 1-го поколения при мейозе образуются 4 типа гамет.
Где гены комбинируются в различных сочетаниях. Такие сочетания получились потому что гены находились в разных хромосомах.
Но в 1906 году Уильям Бэтсон и Риджинальд Пэннет, проводя скрещивание растений душистого горошка и анализируя наследование нескольких признаков формы пыльцы и окраски цветков, обнаружили, что эти признаки не дают независимого распределения в потомстве в соотношении 3:1, гибриды всегда повторяли признаки родительских форм.
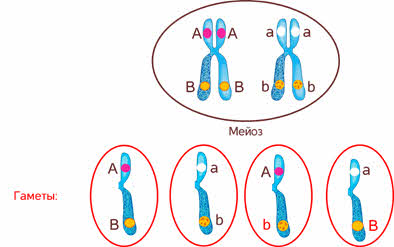
Стало ясно, что не для всех признаков характерно независимое распределение в потомстве и свободное комбинирование.
Дело в том, что ген формы пыльцы и ген окраски цветка лежат в одной хромосоме.
Признаков в организме значительно больше чем хромосом, в которых эти признаки локализованы. Следовательно, каждая хромосома несёт не один ген, а целую группу генов, отвечающих за развитие разных признаков.
Изучением наследования признаков, гены которых локализованы в одной хромосоме, занимался Томас Морган.
Он предложил закон сцепленного наследования (закон Моргана): гены, которые находиться в одной хромосоме, при мейозе попадают в одну гамету, то есть наследуются сцеплено.
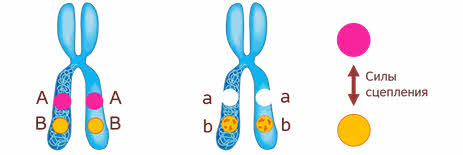
Что значит сцеплено? То есть между генами, которые находиться в одной хромосоме возникают силы сцепления, то есть силы взаимодействия. И чем ближе эти гены, тем сильнее взаимодействие.
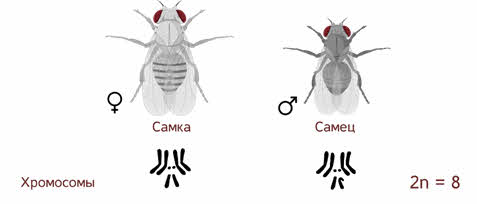
Если Мендель проводил свои опыты на горохе, то для Моргана основным объектом стала фруктовая мушка дрозофила, которая имела диплоидный набор из 8 хромосом.
Небольшие размеры, короткий жизненный цикл и простота культивирования позволяет использовать ряд видов дрозофил как образцовые объекты генетических исследований.
Самец и самка внешне хорошо различимы — у самца брюшко меньше и темнее.
Дрозофила фруктовая — наиболее важный для научных исследований вид дрозофил. Главными её характеристиками как модельного объекта является малое число хромосом. Дрозофила каждые две недели при температуре 25 °С достаточно легко размножаются в пробирках и даёт многочисленное потомство.
Рассмотрим один из первых экспериментов Томаса Моргана по изучению сцепленного наследования.
Скрещивая мушку дрозофилу с серым телом и нормальными крыльями с мушкой, имеющей темную окраску тела и зачаточные крылья, в первом поколении Морган получал гибриды, которые имели серое тело и нормальные крылья.
Ген А-большое отвечает за серое тело, рецессивный ген а-малое за чёрную окраску тела, доминантный ген В-большое за развитие длинных крыльев, а рецессивный ген бэ-малое за не развитие крыльев, то есть крылья остаются в зачаточном состоянии.
Значит ген, определяющий серую окраску брюшка, доминирует над темной окраской, а ген, обуславливающий развитие нормальных крыльев, — над геном недоразвитых.
Далее при скрещивании двух гетерозигот которые имеют серое тело и длинные крылья в первом поколении получается два фенотипических класса организмов.
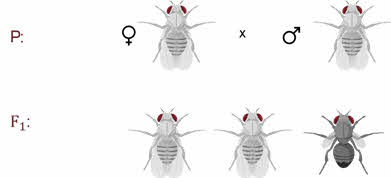
Которые имеют серое тело и длинные крылья и чёрное тело с зачаточными крыльями.
То есть здесь расщепление идёт именно на 2 класса, а не на четыре как при дигибридном скрещивании Менделя.
Почему два? Дело в том, что гены окраски тела и длинны крыльев сцеплены в хромосоме.
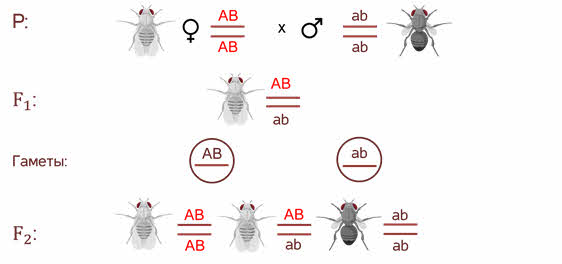
Символы АB АB ab аb располагаются не рядом как мы записывали их ранее, а как бы друг под другом с двумя чёрточками. Чёрточками мы условно обозначаем хромосомы.
В первом поколении организм гетерозиготен по обоим генам, но при образовании гамет эта гетерозиготность не даёт всех возможных комбинаций. То есть родительские гены остаются связанные между собой и гаметы получаются 2х типов.
При комбинации такого рода гамет в потомстве возникают всего 3 генотипических класса потомков.
Морган исследуя наследование сцепленных генов обнаружил что существует нарушение этого правила по дигибридному скрещиванию Менделя.
Он провёл анализирующее скрещивание.
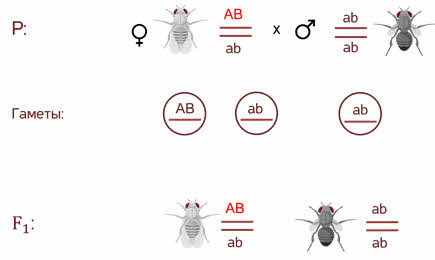
Взял дигетерозиготную особь, которая получилась при скрещивании в первом поколении и скрестил её с чёрной мушкой с зачаточными крыльями, то есть оба рецессивных признака. У него получился необычный результат.
Морган рассуждал. Если гены окраски тела и формы крыльев локализованы в одной хромосоме, то при данном скрещивании должны были получиться две группы особей, повторяющие признаки родительских форм, так как материнский организм должен образовывать гаметы только двух типов — АB и аb, а отцовский — один тип — аb. Следовательно, в потомстве должны образовываться две группы особей, имеющих генотип АB аb и аa BB.
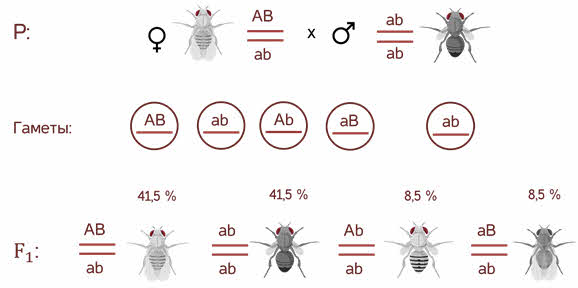
Однако в потомстве появляются особи (пусть и в незначительном количестве) с перекомбинированными признаками, то есть имеющие генотип Аa bb и aa Bb.
В потомстве явно преобладали особи с признаками родительских форм (41,5% были серые длиннокрылые и 41,5% — черные с зачаточными крыльями), и лишь незначительная часть мушек имела иное, чем у родителей, сочетание признаков (8,5% были серые с зачаточными крыльями и 8,5% — черные длиннокрылые).
Такие результаты могли быть получены только в том случае, если гены, отвечающие за окраску тела и форму крыльев, находятся в одной хромосоме.
В профазе первого мейотического деления гомологичные хромосомы (то есть идентичные хромосомы одной пары) конъюгируют (сближаются), и могут разрываться в месте контакта в этот момент между ними может произойти обмен участками – кроссинговер.
Кроссинго́вер или перекрёст — это процесс обмена участками гомологичных хромосом во время конъюгации в профазе первого мейоза. В результате образуются крассоверные гаметы.
Организмы, которые возникают в результате слияния кроссоверных гамет называются рекомбинантными.
Так в результате, кроссинговера в некоторых клетках происходит обмен участками хромосом между генами А и В, появляются гаметы Аb и аB, и, как следствие, в потомстве образуются четыре группы фенотипов, как при свободном комбинировании генов.
Однако кроссинговер происходит не после каждой конъюгации (сближения хромосом). И определить в каких участках хромосом он произойдет достаточно сложно.
В ходе эксперимента Томасу Моргану удалось доказать, что частота кроссинговера между генами прямо пропорциональна расстоянию между ними в хромосоме. То есть можно сказать что, чем дальше гены находятся друг от друга в хромосоме, тем чаще между ними происходит кроссинговер.
Если рассматривать 2 гена А и В мы можем увидеть 2 случая.
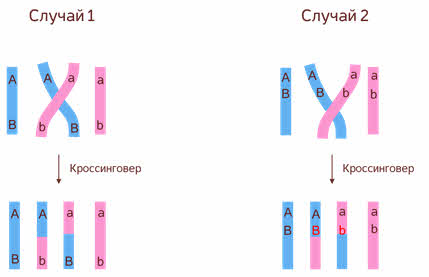
В первом случае гены А и В находиться по разные стороны перекрёста. Тогда после прохождения кроссинговера мы увидим новые сочетания аллелей этих двух генов. В данном случае Аb и аB.
Во 2м случае гены А и B находиться по одну сторону от перекрёста. Тогда после прохождения кроссинговера новых сочетаний аллелей данных двух генов мы не увидим.
Таким образом существуют понятия полного и неполного сцепления.
Неполное сцепление — это разновидность сцепленного наследования, при котором гены анализируемых признаков располагаются на некотором расстоянии друг от друга, что делает возможным кроссинговер между ними.
Полное сцепление — это разновидность сцепленного наследования, при котором гены анализируемых признаков располагаются так близко друг к другу, что кроссинговер между ними становится невозможным.
Это открытие позволило лаборатории Томаса Моргана разработать метод. Который позволяет построить хромосомные карты.
Хромосомные карты — это графическое изображение хромосомы, на котором определенные локусы (гены) отмечены соответственно расстоянию между ними.
Хромосомные карты составляют при помощи генетического анализа, который позволяет точно определить местоположение в хромосоме любого гена.
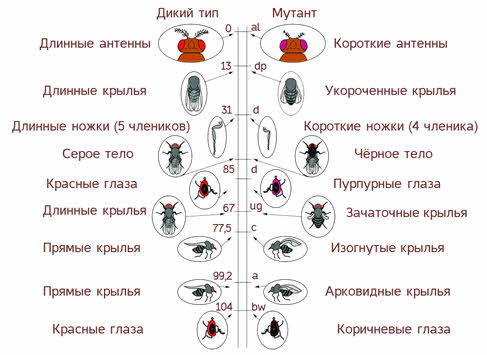
Хромосомная теория наследственности
Изучение Морганом наследования родительских признаков гибридами дрозофилы показало, что число групп сцепленного наследования было равным количеству пар гомологичных хромосом.
Например, у человека 46 хромосом, следовательно, 23 группы сцепления. У дрозофилы 8 хромосом, то есть 4 группы сцепления.
На этом основании был сделан вывод о строгой локализации конкретных генов в определенных парах хромосом.
Возникновение кроссоверных (рекомбинантных) особей дрозофилы можно было объяснить только линейным расположением генов в хромосомах и их обменом при кроссинговере в профазе первого мейоза.
Томас Морган обосновал хромосомную теорию наследственности. Согласно этой теории, передача наследственной информации связана с хромосомами, в которых линейно, в определенной последовательности, локализованы гены. Таким образом, именно хромосомы представляют собой материальную основу наследственности.
Формированию хромосомной теории способствовали данные, полученные при изучении генетики пола, когда были установлены различия в наборе хромосом у организмов различных полов.
Хромосомная теория наследственности сформулирована в 1911 г. американским учёным Томасом Морганом. Её сущность заключается в следующем:
· Основным материальным носителем наследственности являются хромосомы с локализованными в них генами.
· Гены в хромосомах расположены линейно, каждый ген имеет определенное место (локус) в хромосоме;
· Гены, расположенные в одной хромосоме, образуют группу сцепления и наследуются совместно;
· Число групп сцепления равно гаплоидному набору хромосом у гомогаметных особей и n+1 у гетерогаметных особей.
· Между гомологичными хромосомами может происходить обмен участками (кроссинговер); в результате кроссинговера возникают гаметы, хромосомы которых содержат новые комбинации генов.
· Сцепление генов может нарушаться в результате кроссинговера;
· Частота кроссинговера между гомологичными хромосомами зависит от расстояния между генами, локализованными в одной хромосоме. Чем это расстояние больше, тем выше частота кроссинговера.
Значение этой теории заключается в том, что она дала объяснение законам Менделя, вскрыла цитологические основы наследования признаков и генетические основы теории естественного отбора.
Сцепление генов - это совместное наследование генов, расположенных в одной и той же хромосоме. Количество групп сцепления соответствует гаплоидному числу хромосом, то есть у дрозофилы 4; у КРС - 30. Природу сцепленного наследования объяснил в 1910 г. Морган с сотрудниками. В качестве объекта исследования они избрали плодовую муху дрозофилу, которая оказалась очень удобной моделью для изучения данного феномена, так в клетках ее тела, находится только 4 пары хромосом и имеет место высокая скорость плодовитости (в течение года можно исследовать более 20-ти поколений). Итак, сцепленными признаками называются признаки, которые контролируются генами, расположенными в одной хромосоме. Естественно, что они передаются вместе в случаях полного сцепления (закон Моргана).
Полное сцепление встречается редко, обычно – неполное, из-за влияния кроссинговера (перекрещивания и обмена участками гомологичных хромосом в процессе мейоза). То есть, гены одной хромосомы переходят в другую, гомологичную ей.
Частота кроссинговера зависит от расстояния между генами. Чем ближе друг к другу расположены гены в хромосоме, тем сильнее между ними сцепление и тем реже происходит их расхождение при кроссинговере, и, наоборот, чем дальше друг от друга отстоят гены, тем слабее сце пление между ними и тем чаще возможно его нарушение.

На рисунке 1 :
Слева: расстояние между генами А и В маленькое, вероятность разрыва хроматиды именно между А и В невелика, поэтому сцепление полное, хромосомы в гаметах идентичны родительским (два типа), других вариантов не появляется. Справа: расстояние между генами А и В большое, повышается вероятность разрыва хроматиды и последующего воссоединения крест-накрест именно между А и В, поэтому сцепление не
полное, хромосомы в гаметах образуются четырех типов - 2 идентичные родительским (некроссоверные) + 2 кроссоверных варианта.
Количество разных типов гамет бу дет зависеть от частоты кроссинговера или расстояния между анализируемыми генами. Расстояние между генами исчисляется в морганидах: единице расстояния между генами, находящимися в одной хромосоме, соответствует 1% кроссинговера. Такая зависимость между расстояниями и частотой кроссинговера прослеживается только до 50 морганид. Частота кроссинговера между определенной парой генов – довольно постоянная величина (хотя радиация, химические вещества, гормоны, лекарства влияют на нее; например, высокая температура стимулирует кроссинговер).
Пример, основанный на опытах Моргана
Рисунок 2
Фенотипы
А-серое тело, нормальные крылья (повторяет материнскую форму)
Б-тёмное тело, короткие крылья (повторяет отцовскую форму)
В-серое тело, короткие крылья (отличается от родителей)
Г-тёмное тело, нормальные крылья (отличается от родителей)
В и Г получены в результате кроссинговера в мейозе.
Если скрестить мушку дрозофилу, имеющую серое тело и нормальные крылья (на рисунке самка), с мушкой, обладающей тёмной окраской и зачаточными (короткими) крыльями (на рисунке самец), то в первом поколении гибридов все мухи будут серыми с нормальными крыльями (А). Это гетерозиготы по двум парам аллельных генов, причём ген, определяющий серую окраску брюшка, доминирует над тёмной окраской, а ген, обусловливающий развитие нормальных крыльев, - доминирует над геном недоразвитых крыльев.
При анализирующем скрещивании гибрида F1 с гомозиготной рецессивной дрозофилой (Б) подавляющее большинство потомков F2 будет сходно с родительскими формами.
Это происходит потому, что гены, отвечающие за серое тело и нормальные крылья - Сцепленные гены, также как и гены, отвечающие за тёмное тело и короткие крылья, т.е. они находятся в одной хромосоме. наследование сцепленных генов называют - сцепленное наследование.
Сцепление может нарушаться. Это доказывают особи В и Г на рисунке, т. е. если бы сцепление не нарушалось, то этих особей бы не существовало, однако они есть. Это происходит в результате кроссинговера, который и нарушает сцепленность этих генов.
На рисунке 3 опыт Моргана отображен подробно.


Для решения задач важно уловить механизм, поэтому ниже на схемах
еще раз посмотрите, чем отличается независимое (несцепленное) наследование от сцепленного.
Несцепленное наследование: два гена находятся в разных хромосомах, гетерозигота с равной вероятностью дает четыре т ипа гамет:
Сайт учителей биологии МБОУ Лицей № 2 г. Воронежа, РФ
Site biology teachers lyceum № 2 Voronezh city, Russian Federation
В начале XX в., когда генетики стали проводить множество экспериментов по скрещиванию на самых различных объектах (кукуруза, томаты, мыши, мушки дрозофилы, куры и др.), обнаружилось, что не всегда проявляются закономерности, установленные Менделем. Например, не во всех парах аллелей наблюдается доминирование. Вместо него возникают промежуточные генотипы, в которых участвуют обе аллели. Обнаруживается также много пар генов, не подчиняющихся закону независимого наследования генов, особенно если пара аллельных генов находится в одной и той же хромосоме, т. е. гены как бы сцеплены друг с другом. Такие гены стали называть сцепленными.
Механизм наследования сцепленных генов, а также местоположение некоторых сцепленных генов установил американский генетик и эмбриолог Т. Морган. Он показал, что закон независимого наследования, сформулированный Менделем, действителен только в тех случаях, когда гены, несущие независимые признаки, локализованы в разных негомологичных хромосомах. Если же гены находятся в одной и той же хромосоме, то наследование признаков происходит совместно, т. е. сцепленно. Это явление стали называть сцепленным наследованием, а также законом сцепления или законом Моргана.
Закон сцепления гласит: сцепленные гены, расположеные в одной хромосоме, наследуются совместно (сцепленно) .
Примеров сцепленного наследования генов известно очень много. Например, у кукурузы окраска семян и характер их поверхности (гладкие или морщинистые), сцепленные между собой, наследуются совместно. У душистого горошка (Lathyrus odoratus) сцепленно наследуются окраска цветков и форма пыльцы.
Все гены одной хромосомы образуют единый комплекс – группу сцепления. Они обычно попадают в одну половую клетку – гамету и наследуются вместе.
Группа сцепления — все гены одной хромосомы. Число групп сцепления равно количеству хромосом в гаплоидном наборе. Например, у человека 46 хромосом — 23 группы сцепления, у гороха 14 хромосом — 7 групп сцепления, у плодовой мушки дрозофилы 8 хромосом — 4 группы сцепления.
Гены, входящие в группу сцепления, не подчиняются третьему закону Менделя о независимом наследовании. Однако полное сцепление генов встречается редко. Если гены располагаются близко друг к другу, то вероятность перекреста хромосом мала и они могут долго оставаться в одной хромосоме, а потому будут передаваться по наследству вместе. Если же расстояние между двумя генами на хромосоме велико, то существует большая доля вероятности, что они могут разойтись по разным гомологичным хромосомам. В этом случае гены подчиняются закону независимого наследования.
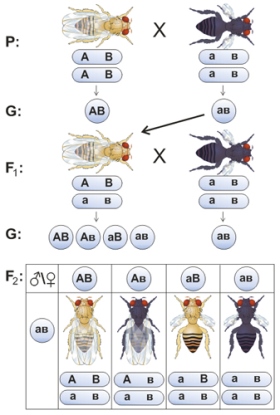
Неполное сцепление генов. При анализе наследования сцепленных генов было обнаружено, что иногда сцепление может нарушаться в результате кроссинговера, происходящего во время мейоза при образовании половых клеток.
Если место разрыва хромосом во время обмена участками расположено между генами А (а) и В(b), то появятся гаметы Ab и аВ , а в потомстве образуются четыре группы фенотипов, как при несцепленном наследовании генов. Отличие заключается в том, что численное соотношение фенотипов не будет соответствовать соотношению 1:1:1:1, как при дигибридном анализирующем скрещивании.
Чем дальше друг от друга находятся гены в хромосоме, тем выше вероятность перекрёста между ними, тем больше процент гамет с перекомбинированными генами, а следовательно, и больше процент особей, отличных от родителей. Такое явление называют неполным сцеплением генов .
На рисунке - Наследование при неполном сцеплении генов (на примере скрещивания двух линий дрозофил, где А – нормальные крылья, а – зачаточные крылья, В – серый цвет тела, в – черный цвет тела ).
Полное сцепление генов. Чем ближе друг к другу находятся гены в хромосоме, тем меньше вероятность перекрёста между ними. Если гены расположены очень близко друг к другу (рядом), то перекрёста между ними обычно не наблюдается. В этом случае говорят о полном сцеплении генов .
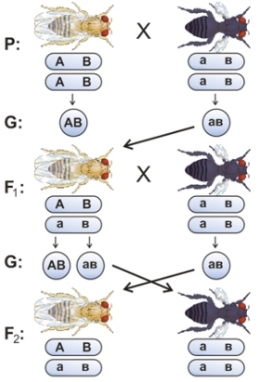
На рисунке - Наследование с полным сцеплением генов (на примере скрещивания двух линий дрозофил, где А – нормальные крылья, а – зачаточные крылья, В – серый цвет тела, в – черный цвет тела )
Таким образом, третий закон Менделя отражает частое, но не абсолютное явление в наследовании признаков.
Основные доказательства передачи наслед-ственности были получены в экспериментах Моргана и его сотрудников.
Таким образом, сцепленное наследование — явление совместного наследования генов, локализованных в одной хромосоме.
Читайте также:
- Как называется кредитная организация функция которой обеспечить расчеты на бирже
- Как уволиться если не отпускают
- Можно ли штрафовать инвалида 2 группы за административное правонарушение
- За какие деяния по вопросам призыва гражданин может быть привлечен к уголовной ответственности
- 52 чему равна полная и полезная мощность а также кпд в режиме согласования