
Что осуществляет избирательный транспорт веществ в клетке
Обновлено: 30.06.2024
Транспортные белки – это белки, которые транспортируют вещества через биологические мембраны. Транспортные белки находятся внутри самой мембраны, где они образуют канал или несущий механизм, чтобы позволить их подложка переходить с одной стороны на другую.
Вещества, переносимые этими белками, могут включать ионы, такие как натрий и калий; сахара, такие как глюкоза; белки и молекулы-мессенджеры; и многое другое.
Транспортные белки обычно осуществляют два вида транспорта:облегченная диффузия ”, Где транспортный белок просто создает отверстие для вещества, чтобы распространять вниз его градиент концентрации ; а также “активный транспорт,” где клетка расходует энергию, чтобы переместить вещество против градиента его концентрации.
Функция транспортного белка
Жизнь, как мы знаем, зависит от способности клеток избирательно перемещать вещества, когда им это необходимо. Определенные важные молекулы, такие как ДНК, должны постоянно находиться внутри клетки; но другие молекулы, такие как ионы, сахара и белки, возможно, должны пройти внутрь и наружу, чтобы клетка функционировала должным образом.
Перемещая вещества через мембраны, транспортные белки делают возможным все – от нервных импульсов до клеточного метаболизма.
Без транспортных белков, например, градиент натрия-калия, который позволяет нашим нервам срабатывать, не существовал бы.
Типы транспортных белков
Каналы / Поры
Эти белки характеризуются тем, что они открыты как для внутриклеточного, так и для внеклеточного пространства одновременно. Напротив, белки-носители открыты только внутри или снаружи клетки в любой момент времени.
Каналы или поры обычно сконструированы так, что через них может проходить только одно конкретное вещество.
Например, в ионно-управляемых каналах напряжения часто используются заряженные аминокислоты, расположенные на точных расстояниях, чтобы привлечь их желаемый ион, отталкивая все остальные. Затем желаемый ион может течь через канал, тогда как другие вещества не могут.
Ионные каналы, управляемые напряжением, являются хорошими примерами транспортных белков, которые действуют по мере необходимости. Часто обнаруживаемые в нейронах, управляемые напряжением ионные каналы открываются в ответ на изменения электрохимического потенциала мембраны.
В закрытом состоянии управляемый напряжением канал не позволяет ионам проходить через клеточная мембрана, Но когда он открыт, он позволяет огромным количествам ионов проходить очень быстро, позволяя клетке быстро менять свой мембранный потенциал и запускать нервный импульс.
Белки-носители
Белки-носители представляют собой транспортные белки, которые открыты только с одной стороны мембраны одновременно.
Они часто проектируются таким образом, потому что они транспортируют вещества против градиента их концентрации. Одновременное открывание по обе стороны мембраны может позволить этим веществам просто течь обратно вдоль градиента концентрации, сводя на нет белок-носитель Работа
Для выполнения своей работы белки-носители обычно используют энергию для изменения формы.
Например, натриево-калиевая помпа использует энергию АТФ, чтобы изменить свою форму с открытой на внутриклеточную. решение, чтобы быть открытым для внеклеточного раствора. Это позволяет ему собирать ионы внутри клетки и выпускать их снаружи, а затем наоборот.
Другие белки-носители могут использовать другие источники энергии, такие как существующие градиенты концентрации, для достижения «вторичного активный транспорт «. Это означает, что их транспорт стал возможен благодаря энергии, расходуемой клеткой, но сам белок не использует АТФ напрямую.
Хорошим примером является транспортный белок натрий-глюкоза, который использует градиент концентрации натрия, первоначально созданный натриево-калиевым насосом, для перемещения глюкозы против градиента концентрации.
Мы обсуждаем натрий-калиевый насос и натрий-глюкозный транспортный белок подробно ниже.
Примеры транспортных белков
Натриево-калиевый насос
Наиболее известным примером первичного активного транспортного белка является натриево-калиевая помпа. Именно этот насос создает ионный градиент, который позволяет нейронам срабатывать.
Натриево-калиевая помпа начинается с натриевых сайтов связывания, обращенных внутрь клетки. Эти сайты привлекают ионы натрия и удерживают их.
Когда каждый из трех сайтов связывания натрия связывает ион натрия, белок связывается с молекула АТФ, и разбивает его на ADP + фосфатная группа, Белок использует энергию, выделяемую в этом процессе, чтобы изменить форму.
Теперь сайты связывания натрия сталкиваются с внеклеточным раствором. Они выделяют три иона натрия вне клетки, в то время как сайты связывания белка с калием связываются с двумя ионами калия.
Когда оба калий-связывающих сайта заполнены, белок возвращается к своей первоначальной форме. Теперь ионы калия высвобождаются внутри клетки, и пустые сайты связывания натрия могут связывать больше ионов натрия.

Когда наступает время для срабатывания нервной клетки, сильные электрические и химические градиенты позволяют клетке производить огромное мгновенное изменение, открывая свои управляемые напряжением ионные каналы.
Натрий-глюкоза транспортные белки
Белок транспорта натрия-глюкозы использует вторичный активный транспорт для перемещения глюкозы в клетки. Они активны в клетках кишечника и почек, которые должны перемещать глюкозу в системы организма против градиента концентрации.
Эта операция требует энергии, потому что рассматриваемые клетки имеют более высокую концентрацию глюкозы, чем внеклеточной жидкости, Следовательно, глюкоза не сможет самостоятельно диффундировать в клетки; энергия должна быть приложена.
В этом случае энергия поступает из градиента концентрации натрия. Благодаря действию натриево-калиевого насоса, натрий находится вне клетки гораздо больше, чем внутри нее. Таким образом, существует сильный градиент концентрации, способствующий движению натрия в клетку.
Закрытые ионные каналы в улитке
Закрытые ионные каналы пассивный транспорт белки, которые открываются в ответ на конкретные раздражители. Возможно, вы знакомы с ионно-управляемыми ионными каналами, такими как те, которые вызывают срабатывание наших нейронов в ответ на сигналы, поступающие от других нейронов.
Менее известными являются стробированные ионные каналы улитки, которые открываются механическим давлением вместо изменений напряжения. Эти замечательные ионные каналы позволяют нервам нашего внутреннего уха срабатывать в ответ на вибрации звука. Вот как мы слышим.
Внутренние волосковые клетки, с другой стороны, имеют особую работу. В ответ на эти вибрации они открывают ионные каналы в своих клеточных мембранах и освобождают нейротрансмиттеры, как это делает нейрон.
Эти нейротрансмиттеры вызывают укол соседних нервов. И вот как звук преобразуется в нервные импульсы!
- Активный транспорт – Транспорт веществ через биологические мембраны, что требует от клетки расходовать энергию.
- Закрытый ионный канал – Белок, который позволяет ионам проходить, когда открыт, который открывается в ответ на определенный стимул.
- Пассивный транспорт – Транспорт веществ через биологические мембраны, который происходит естественным путем, без необходимости тратить энергию.
викторина
Ответ на вопрос № 1
2. В чем разница между первичным и вторичным активным транспортом?A. Первичный активный транспорт использует белки-носители, тогда как вторичный активный транспорт использует белки-каналы.B. Первичный активный транспорт может транспортировать только одно вещество за раз, тогда как вторичный активный транспорт может транспортировать два.C. Первичный активный транспорт требует энергии; вторичного активного транспорта нет.D. Первичные активные транспортные белки используют АТФ напрямую. Вторичные активные транспортные белки используют энергию, которая получается из других АТФ-зависимых процессов.
Ответ на вопрос № 2
D верно. Все виды активного транспорта требуют от клетки расходовать энергию. Первичные активные транспортные белки берут энергию непосредственно из АТФ; вторичные активные транспортные белки используют энергию от процессов, происходящих из АТФ.
3. Что из перечисленного НЕ является примером активного транспорта?A. Натриево-калиевый насос перемещает натрий и калий как против градиента их концентрации.B. Ионные каналы волосковых клеток открываются в ответ на давление, позволяя ионам проходить через них.C. Транспортер натрия-глюкозы использует градиент концентрации натрия для перемещения глюкозы в клетку.D. Ни один из вышеперечисленных.
Ответ на вопрос № 3
В верно. Ионные каналы являются формой пассивного транспорта; они позволяют ионам двигаться вниз по градиенту концентрации, что не требует затрат энергии.
Оболочки клеток всех организмов содержат плазматическую мембрану. Мембрана клетки (а так же всех мембранных органоидов) состоит из белков и (фосфо)липидов.
У животных клеток наружный слой мембраны, состоящий из углеводов, присоединенных к белкам и липидам мембраны, называется гликокаликс.
У всех, кроме животных, снаружи от мембраны имеется твердая клеточная стенка. У растений она состоит из целлюлозы (клетчатки), у грибов из хитина, у бактерий из муреина. Клеточная стенка обеспечивает защиту клетки, опору и тургор (вода за счет осмоса надувает клетку изнутри, клеточная стенка напрягается, клетка становится упругой).
Функции плазматической мембраны
1) Избирательная проницаемость: мембрана регулирует обмен веществ между клеткой и окружающей средой. Регуляцию осуществляют мембранные белки:
- каждый из них переносит только одно определенное вещество (специфичность),
- при изменении условий белок может изменить (прекратить) свою работу (денатурация).
2) Сигнальная функция: в мембрану встроены белки, которые при изменении условий снаружи клетки или при присоединении какого-либо сигнального вещества обратимо денатурируют и передают сигнал в клетку.
3) Фагоцитоз: захват и поглощение животной клеткой крупных частиц (открыл И.И.Мечников): в том месте, где поверхность клетки соприкасается с частицей, образуется углубление, мембрана окружает частицу со всех сторон. Затем часть мембраны отделяется и внутри цитоплазмы оказывается фагоцитозный пузырек с веществом внутри. Например: амеба поглощает пищу, фагоцит поглощает чужеродную частицу. (Пиноцитоз – захват и поглощение капель жидкости.)
Транспорт веществ через мембрану
1) Активный транспорт (идёт с затратой энергии АТФ). Например, белок-канал натрий-калиевая АТФ-аза выкачивает из клетки ионы натрия и закачивает ионы калия.
2) Пассивный транспорт (диффузия, идет без затрат энергии). Например, газы воздуха и вода могут проходить непосредственно между фосфолипидами.
К пассивному транспорту относится осмос, когда вода движется через мембрану в область большей концентрации веществ. Из-за осмоса эритроцит в пресной воде лопается (в него заходит слишком много воды), а в пересолёной воде – сморщивается (вода выходит).
Еще можно почитать
Задания части 1
Выберите один, наиболее правильный вариант. В процессе обмена веществ в клетке энергия АТФ может использоваться
1) для выделения углекислого газа из клетки
2) для образования воды на кислородном этапе энергетического обмена
3) при расщеплении биополимеров
4) на поступление веществ в клетку через плазматическую мембрану
Выберите один, наиболее правильный вариант. Активный транспорт веществ в клетку и из клетки из области с низкой концентрацией в область с высокой концентрацией происходит
1) без затраты энергии
2) путем фагоцитоза
3) путем осмоса
4) с затратами энергии
Выберите один, наиболее правильный вариант. Эритроциты, помещенные в физиологический раствор
1) набухают и лопаются
2) остаются без внешних изменений
3) сморщиваются
4) слипаются друг с другом
Выберите один, наиболее правильный вариант. Поступление воды в растительную клетку происходит в процессе
1) осмоса
2) фагоцитоза
3) пиноцитоза
4) активного транспорта
Выберите один, наиболее правильный вариант. Мембраны хлоропластов образованы молекулами
1) целлюлозы
2) хитина
3) гликогена
4) белков и липидов
Выберите один, наиболее правильный вариант. Плазматическая мембрана животной клетки в отличие от клеточной стенки растений
1) состоит из клетчатки
2) состоит из белков и липидов
3) прочная, неэластичная
4) проницаема для всех веществ
Выберите один, наиболее правильный вариант. Поступление питательных веществ путем фагоцитоза происходит в клетках
1) прокариот
2) животных
3) грибов
4) растений
2. Выберите три верных ответа из шести и запишите цифры, под которыми они указаны. Плазматическая мембрана животной клетки состоит из
1) митохондрий
2) рибосом
3) двух слоев липидов
4) молекул белков
5) двух слоев углеводов
6) углеводов, образующих гликокаликс
Установите последовательность увеличения уровня организации структур клетки, начиная с наименьшей. Запишите в таблицу соответствующую последовательность цифр.
1) молекула глицерина
2) липидный бислой
3) фосфолипид
4) цитоплазма клетки
5) мембрана ЭПС
МЕМБРАНА ФУНКЦИИ
1. Выберите три варианта. Какие функции выполняет в клетке плазматическая мембрана?
1) придает клетке жесткую форму
2) отграничивает цитоплазму от окружающей среды
3) служит матрицей для синтеза иРНК
4) обеспечивает поступление в клетку ионов и мелких молекул
5) обеспечивает передвижение веществ в клетке
6) участвует в поглощении веществ клеткой
2. Выберите три правильных варианта. Какие из перечисленных функций выполняет плазматическая мембрана клетки?
1) участвует в синтезе липидов
2) осуществляет активный транспорт веществ
3) участвует в процессе фагоцитоза
4) участвует в процессе пиноцитоза
5) является местом синтеза мембранных белков
6) координирует процесс деления клетки
МЕМБРАНА - СТЕНКА
Установите соответствие между функциями и структурами клетки: 1) клеточная мембрана, 2) клеточная стенка. Запишите цифры 1 и 2 в порядке, соответствующем буквам.
А) избирательная проницаемость
Б) активный транспорт
В) поддержание формы клетки
Г) придает жесткость клетки
Д) способность к фагоцитозу

МЕМБРАНА - АГ
Установите соответствие между функциями клеточных структур и структурами, изображёнными на рисунке. Запишите цифры 1 и 2 в порядке, соответствующем буквам.
А) осуществляет активный транспорт веществ
Б) изолирует клетку от окружающей среды
В) обладает избирательной проницаемостью
Г) образует секреторные пузырьки
Д) распределяет вещества клетки по органеллам
Е) участвует в образовании лизосом
ФАГОЦИТОЗ
Установите соответствие между клетками и способностью к фагоцитозу: 1) способны 2) не способны. Запишите цифры 1 и 2 в правильном порядке.
А) амёба
Б) лейкоциты
В) хлорелла
Г) мукор
Д) инфузория
Е) хламидомонада
1. Установите последовательность процессов, происходящих в клетке амёбы при питании. Запишите соответствующую последовательность цифр.
1) слияние фагоцитозного пузырька с лизосомой
2) впячивание плазматической мембраны с частицами пищи
3) отделение мембранного пузырька с частицей пищи внутрь клетки
4) замыкание плазматической мембраны с образованием фагоцитозного пузырька
5) расщепление биополимеров пищи до мономеров
2. Установите последовательность процесса фагоцитоза. Запишите соответствующую последовательность цифр.
1) гидролитическое расщепление веществ под действием ферментов
2) образование фагоцитозного пузырька и продвижение его внутрь клетки
3) слияние лизосомы с фагоцитозным пузырьком
4) обволакивание плазматической мембраной пищевой частицы
5) поступление мономеров в цитоплазму клетки
3. Установите последовательность процессов, происходящих при фагоцитозе
1) поступление мономеров в цитоплазму
2) захват клеточной мембраной питательных веществ
3) гидролиз полимеров до мономеров
4) образование фагоцитозного пузырька внутри клетки
5) слияние фагоцитозного пузырька с лизосомой

3. Все перечисленные ниже признаки, кроме двух, используются для описания изображенной на рисунке структуры клетки. Определите два признака, выпадающих из общего списка, и запишите цифры, под которыми они указаны.
1) состоит из двух слоев белков и липидов между ними
2) в состав входят фосфолипиды
3) холестерин придает прочность
4) липиды осуществляют транспортную функцию
5) не пропускает крупные и заряженные молекулы
Жизнедеятельность клетки связана с постоянным обменом ее содержимого с окружающей средой. Точно так же и внутри клетки происходит перемещение веществ между органеллами или компартментами. Все эти события связаны с преодолением основного барьера для веществ — мембраны, ограничивающей органеллу или саму клетку. При этом следует помнить, что главная функция биомембран — избирательность транспорта для различных - веществ и ионов.
Возможные способы транспорта через мембраны можно разделить на 4 основных типа: пассивная диффузия, облегченная диффузия, активный транспорт и цитозы.
Пассивная диффузия. Это процесс транспорта через мембраны веществ из области с большей их концентрацией в область с меньшей концентрацией (по химическому градиенту), в котором не принимают участия транспортные белки и не затрачивается энергия. С помощью такого способа через мембрану транспортируются малые незаряженные молекулы, например молекулы газов, некоторые анестезирующие вещества, а также вода. Чтобы пересечь бислой, молекула должна преодолеть поверхностное натяжение на границе мембраны, проникнуть в бислой, продиффундировать через него и выйти с противоположной стороны, вновь преодолев энергетический барьер на границе раздела фаз. Этим и объясняется избирательная проницаемость липидного бислоя для небольших молекул неэлектролитов. Удивительным является факт весьма легкого и быстрого проникновения воды через мембраны: показано, что молекуле воды требуется для пересечения бислоя всего 1 мкс. Для объяснения этого феномена в последнее время появляются основанные на некоторых экспериментальных данных предположения о том, что в мембранах все же существуют какие-то белковые проводящие пути для воды, либо молекулы воды пользуются локальными дефектами в структуре бислоев.
Перемещение одних только молекул воды через полупроницаемую мембрану можно рассматривать как частный вид диффузии — осмос. Под осмосом понимают переход молекул воды из области с высоким водным потенциалом и низкой концентрацией растворенного вещества в область с низким водным потенциалом и высокой концентрацией растворенного вещества (рис. 4.9). В этом случае молекулы воды будут переходить из гипотонического раствора в гипертонический до тех пор, пока не наступит равновесие и оба раствора не станут изотоническими по отношению друг к другу.
Перемещение воды через плазматические мембраны клеток в соответствии с законами осмоса создает организмам немалые проблемы, особенно для водных обитателей. Поэтому осморегуляция (поддержание водного потенциала в клетке на постоянном уровне) является важной стороной функциональной деятельности большинства организмов, и на ее осуществление зачастую тратится значительная доля запасенной клеткой энергии.

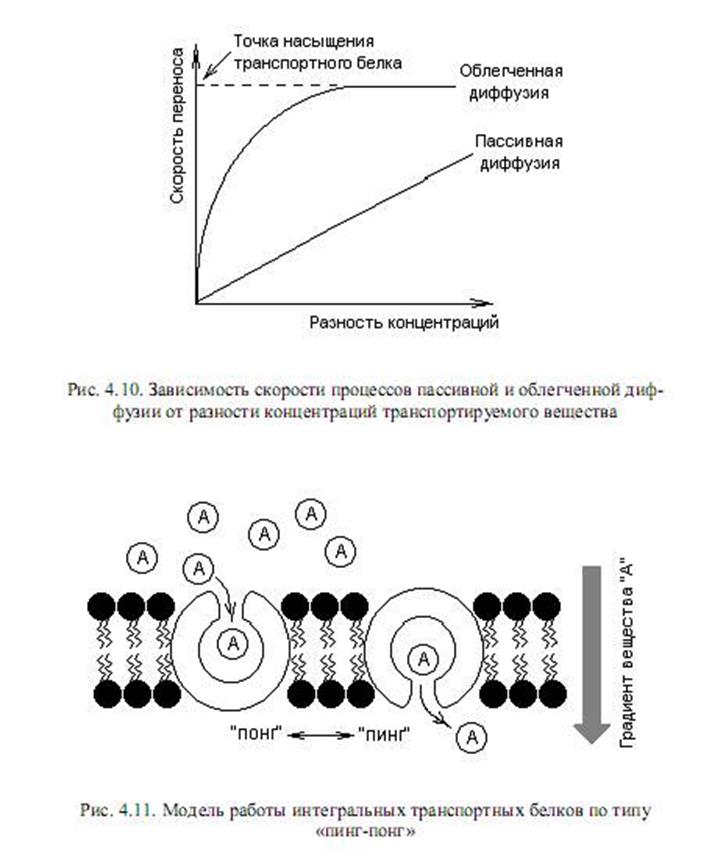
Скорость диффузии веществ через мембраны зависит от многих причин: растворимости вещества в мембране, коэффициента диффузии в мембране, а также разности концентрации вещества снаружи и внутри клетки (градиента концентрации) (рис. 4.10).
Облегченная диффузия. Этот вид транспорта осуществляется с помощью транспортных белков по электрохимическому градиенту (разность электрических потенциалов и концентраций веществ) без затрат энергии. Это селективный перенос веществ — вещество будет транспортировано через мембрану лишь в том случае, если для него в мембране имеется функционирующий транспортный белок.
Поскольку в облегченной диффузии задействованы белки, этот процесс, в отличие от пассивной диффузии, может достигать эффекта насыщения. Стадия насыщения (рис. 4.10) характеризует состояние, когда все транспортные белки для данного вещества насыщены субстратом и скорость транспорта этого вещества достигает максимума.
Работу каналов можно рассмотреть на примере ацетилхолинового рецептора. Этот интегральный белок находится в основном в мембранах нервномышечных соеди-нений скелетных мышц. Он состоит из пяти субъединиц четырех типов и открывается в ответ на связывание ацетилхолина (нейромедиатор). При взаимодействии с ацетилхолином канал открывается, что связано с изменением конформации субъединиц, и пропускает определенные ионы (Na+ , K+ , Ca2+ и некоторые другие), остается в таком положении 1 мс, а затем закрывается.
Селективное перемещение катионов изменяет трансмембранный потенциал, в результате чего происходит электрическое возбуждение мышечной клетки, что приводит к сокращению мышцы. Изучение структуры ацетилхолинового рецептора показало, что пять белковых субъединиц встроены в бислой определенным образом: они организованы вокруг центральной поры диаметром 3 нм, через которую и транспортируются катионы. Непроницаемость канала для анионов и в три раза большую проницаемость для катионов, чем для незаряженных молекул, можно объяснить электростатическими взаимодействиями, возникающими благодаря присутствию в воротах канала биполярных или отрицательно заряженных групп.
Особым типом транспорта веществ в ходе облегченной диффузии является использование ионофоров, действие которых изучено на искусственных мембранах.
Под ионофорами понимают низкомолекулярные вещества пептидной природы, избирательно транспортирующие через мембраны ионы. Различают ионофорыканалообразователи (грамицидин А, амфотерицин B и др.) и ионофорыпереносчики (валиномицин, энниатины, боверицин).
В биологических мембранах ионофорный тип транспорта до сих пор не обнаружен.
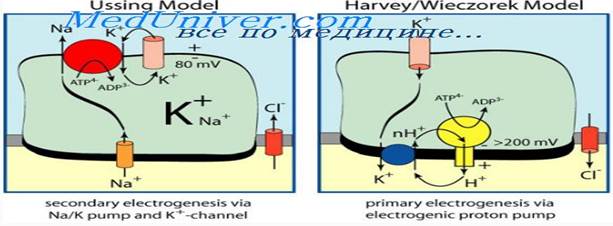
Активный транспорт. Это сопряженный с потреблением энергии перенос молекул или ионов через мембрану против электрохимического градиента, в котором задействованы транспортные белки. Благодаря активному транспорту в жизнеспособных клетках между двумя сторонами мембраны поддерживается разность потенциалов, т. е. электрический заряд, при этом у большинства изученных клеток внутреннее содержимое заряжено отрицательно по отношению к внешней среде.
Активный транспорт сопряжен со значительными затратами энергии: некоторые клетки тратят более трети всей запасенной энергии для создания ионного градиента на мембране. Это необходимо для таких жизненно важных процессов, как осморегуляция, генерация и передача нервных импульсов, перенос в клетки питательных веществ (сахаров, аминокислот и др.).
Разнообразные системы активного транспорта отличаются друг от друга, в первую очередь тем, что служит для них источником энергии: АТР, ионный градиент, фосфоенолпируват, видимый свет. Наиболее хорошо изученной системой активного транспорта является натрийкалиевая (Na+/K+)АТРаза, функционирующая в плазматических мембранах животных клеток. Этот интегральный белок состоит из двух субъединиц: бульшая представлена полипептидом, имеющим участки связывания для ионов натрия и АТР на цито плазматической поверхности, а ионов калия — на наружной; меньшая субъединица является гликопротеином. Работа (Na+/K+)АТРазы заключается в следующем: при гидролизе одной молекулы АТР из клетки выкачивается 3 иона Na+ , а извне в клетку проводится 2 иона K+ , т. е. выводится больше положительных ионов, чем проводится внутрь клетки. Так на внутренней стороне мембраны создается избыточный отрицательный заряд, и клетка становится электрогенной. В мембранах обычно присутствуют проводящие пути для облегченной диффузии ионов натрия и калия по электрохимическому градиенту, и этот транспорт, хотя и с малой скоростью, со-вершается. Однако в жизнеспособной клетке не происходит уравнивания концентраций ионов, создающих электрохимический градиент на мембране, благодаря постоянной работе первичных активных переносчиков, таких, как (Na+/K+)АТРаза. Таким образом, ионные насосы, принимающие участие в первичном активном транспорте, осуществляют перемещение заряда на мембране и создают на ней электрохимический градиент, в котором заключена энергия.
Вторичные активные переносчики используют электрохимические градиенты в качестве движущей силы для транспорта растворимых веществ. Этот процесс можно проследить на примере клеток эпителия кишечника. Образуемые в кишечнике при переваривании пищи строительные блоки (аминокислоты, глюкоза и др.) поступают в кровь при диффузии через мембраны кровеносных сосудов, и эта диффузия осуществляется в ходе симпорта(однонаправленного транспорта) с ионами натрия.
Ионы натрия стремятся возвратиться в клетку согласно закономерностям облегченной диффузии и как бы тянут с собой молекулы питательных веществ. В мембранах обнаружены специфические переносчики сахаров и разных аминокислот, которые функционируют в системе активного транспорта, накапливая в клетке эти вещества, извлекая их даже из очень разбавленных растворов, т. е. против химического градиента. Эти же транспортные системы могут участвовать и в облегченной диффузии, если вещества транспортируются по химическому градиенту. Кроме описанного выше примера симпорта питательных веществ вместе с возвращающимися в клетку ионами натрия, существует и разнонаправленный транспорт — антипорт. Например, белок полосы 3 эритроцитов осуществляет сопряженный транспорт Cl и HCO3 в противоположных направлениях через эритроцитарную мембрану.
У аэробных бактерий транспорт питательных веществ в клетку осуществляется в ходе симпорта не с ионами Na+ , а с протонами. Наилучшим образом охарактеризованным примером подобного переносчика служит лактозопермеаза кишечной палочки. Этот интегральный белок использует протонный электрохимический градиент, созданный на мембране в результате окислительного фосфорилирования, для симпорта лактозы: с каждым возвращенным в клетку протоном переносится одна молекула лактозы.
Следует отметить, что АТРазы представляют собой ферменты, катализирующие взаимообратимые реакции: при гидролизе АТР ионы транспортируются против электрохимического градиента, а перенос ионов по электрохимическому градиенту через каналы АТРазы может запускать синтез АТР.
Эндоцитоз и экзоцитоз. Эти способы переноса веществ через мембраны связаны с образованием впячиваний (инвагинаций) мембраны и формированием особых мембранных везикул, обеспечивающих прохождение через мембрану крупных макромолекул и частиц. При этом эндоцитоз обеспечивает поглощение клеткой веществ, а экзоцитоз — выделение из клетки. Принято делить цитозы еще на два типа: пиноцитоз и фагоцитоз. Пиноцитоз — это механизм, с помощью которого через мембрану проводятся белки и другие макромолекулы в жидкой фазе.
Фагоцитоз представляет собой поглощение клеткой крупных частиц, например бактерий, вирусов. Эти виды транспорта характерны в основном для эукариотических клеток, причем у животных фагоцитоз осуществляют только специализированные клетки, такие, например, как макрофаги. Для многих простейших, например амеб, фагоцитоз является основным способом питания.
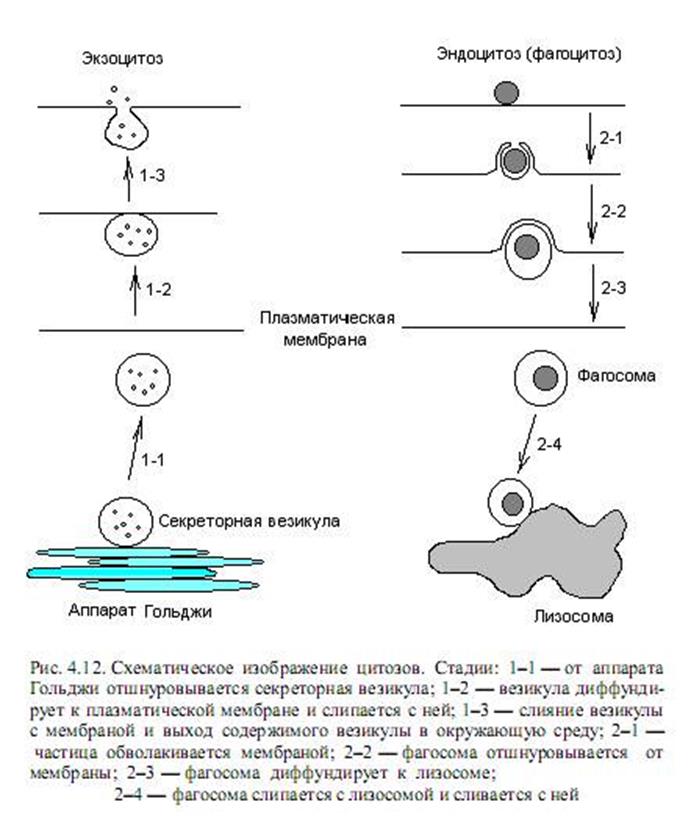
Важной особенностью цитозов является последовательное образование и слияние везикул, в которых заключено транспортируемое вещество, причем секретируемые и поглащаемые молекулы локализуются в везикулах и не смешиваются с другими макромолекулами или органеллами клетки. С помощью не установленного пока меха-низма каждый пузырек сливается только со специфическими мембранными структурами (рис. 4.12).
В основе цитозов лежит еще одно характерное свойство липидных слоев биомембран — способность к агрегации, в результате чего мелкие везикулы объединяются в более крупные или происходит объединение везикул с плазматической мембраной клетки. Такой механизм основан на универсальности структуры биомембран, участвующих в формировании клеточных органелл и протопластов. Аналогичное явление можно наблюдать в пенах, где мыльные пузыри, также состоящие из амфифильных молекул (мыла — соли жирных кислот), обладают тенденцией к объединению с образованием более крупных структур.
Способность мембран к агрегации лежит в основе такого широко распространенного способа переноса генетической информации, как слияние протопластов.

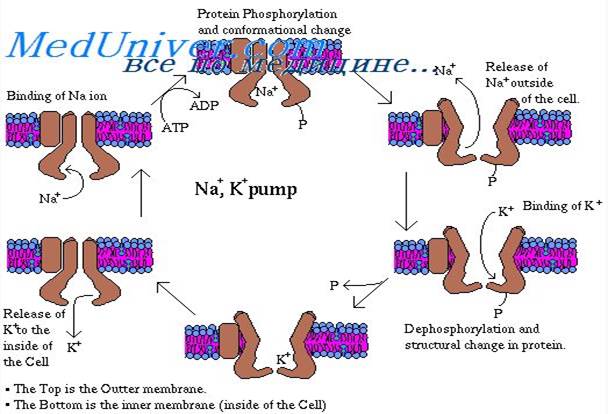
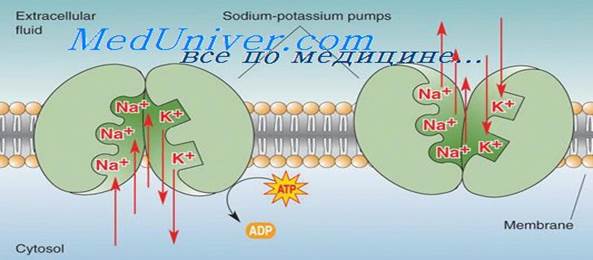
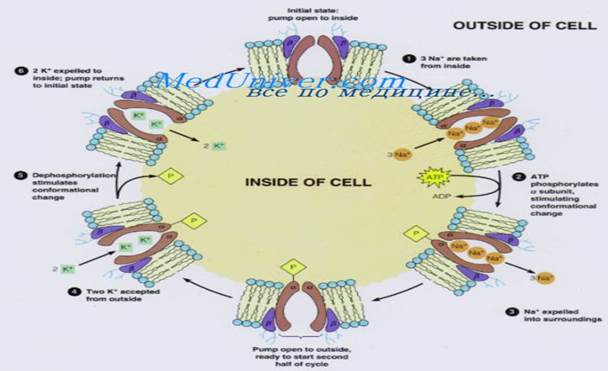
Натрий-калиевый насос. К веществам, которые транспортируются посредством первично активного транспорта, относят натрий, калий, кальций, водород, хлор и некоторые другие ионы. Механизм активного транспорта лучше всего изучен для натрий-калиевого насоса (Na+/K+-нaсоса) — транспортного процесса, который выкачивает ионы натрия через мембрану клетки наружу и в то же время закачивает в клетку ионы калия. Этот насос отвечает за поддержание различной концентрации ионов натрия и калия по обе стороны мембраны, а также за наличие отрицательного электрического потенциала внутри клеток. (В главе 5 будет показано, что он является также основой процесса передачи импульсов в нервной системе.)
Белок-переносчик представлен комплексом из двух раздельных глобулярных белков: более крупного, называемого альфа-субъединицей, с молекулярной массой около 100000, и меньшего, называемого бета-субъединицей, с молекулярной массой около 55000. Хотя функция меньшего белка неизвестна (за исключением того, что он, возможно, закрепляет белковый комплекс в липидной мембране), крупный белок имеет три специфических характеристики, важные для функционирования насоса. 1. На выступающей внутрь клетки части белка имеются три рецепторных участка для связывания ионов натрия. 2. На наружной части белка располагаются два рецепторных участка для связывания ионов калия. 3. Внутренняя часть белка, расположенная вблизи участков связывания ионов натрия, обладает АТФ-азной активностью. Рассмотрим работу насоса. Когда 2 иона калия связываются с белком-переносчиком снаружи и 3 иона натрия связываются с ним внутри, активируется АТФ-азная функция белка. Это ведет к расщеплению 1 молекулы АТФ до АДФ с выделением энергии высокоэнергетической фосфатной связи. Полагают, что эта освобожденная энергия вызывает химическое и конформационное изменение молекулы белка-переносчика, в результате 3 иона натрия перемещаются наружу, а 2 иона калия — внутрь клетки. Как и другие ферменты, Na-K+-ATФ-aзa может работать и в обратном направлении. При экспериментальном увеличении электрохимических градиентов для Na+ и К+ до таких значений, что накопленная в них энергия станет выше химической энергии гидролиза АТФ, эти ионы будут двигаться по своим градиентам концентрации, а Na+/K+-Hacoc будет синтезировать АТФ из АДФ и фосфата. Следовательно, фосфорилированная форма Nа+/К+ -насоса может быть или донором фосфатов для синтеза АТФ из АДФ, или использовать энергию для изменения своей конформации и качать натрий из клетки, а калий — в клетку. Относительные концентрации АТФ, АДФ и фосфатов, как и электрохимические градиенты для натрия и калия, определяют направление ферментативной реакции. Для некоторых клеток, например, электрически активных нервных клеток, от 60 до 70% всей потребляемой клеткой энергии тратится на перемещение натрия наружу и калия внутрь. Роль Na-K-насоса: Активный транспорт ионов кальция и водорода в клетке. Одной из многочисленных важных функций Na+/K+-Hacoca является регуляция объема каждой клетки. Без функционирования этого насоса большинство клеток организма будут набухать, пока не лопнут. Механизм регуляции объема следующий: внутри клетки много белков и других органических молекул, которые не могут покинуть клетку. Большинство из них отрицательно заряжены и потому связывают большое число ионов калия, натрия и других положительно заряженных ионов. Все эти молекулы и ионы вызывают осмос воды в клетку. Без регуляции осмоса клетка будет неограниченно разбухать вплоть до разрыва мембраны. В норме механизмом для предупреждения этого является Na+/K+-Hacoc. Вспомним, что в результате работы насоса 3 иона натрия выводятся наружу, а 2 иона калия закачиваются внутрь. Кроме того, мембрана гораздо менее проницаема для ионов натрия, чем для калия, поэтому ионы натрия, оказавшись снаружи, в основном там и остаются. Следовательно, присутствует общая потеря ионов клеткой, что, в свою очередь, инициирует осмос воды из клетки. Когда клетка начинает разбухать, это автоматически активирует Na+/K+-Hacoc, обеспечивая удаление из клетки еще большего числа ионов вместе с водой. Таким образом, Na+/K+-Hacoc осуществляет непрерывную регуляцию объема клетки, поддерживая его в нормальных пределах. Электрогенная природа натрий-калиевого насоса. Как известно, Na+/K+-Hacoc выкачивает 3 иона натрия наружу на каждые 2 иона калия, входящие внутрь. Это означает, что 1 положительный заряд выводится наружу при каждом цикле работы насоса. Создается избыток положительных зарядов на поверхности клетки и дефицит положительных ионов внутри клетки, т.е. внутренняя часть клетки заряжается отрицательно. В связи с этим Na+/K+-Hacoc называют электрогенным, поскольку он создает трансмембранную разность потенциалов, а наличие электрического потенциала является основой для передачи сигналов в нервных и мышечных волокнах. Первично активный транспорт ионов кальция. Другим важным механизмом первично активного транспорта является кальциевый насос. В норме ионы кальция во внутриклеточном цитозоле практически всех клеток содержатся в чрезвычайно низкой концентрации — примерно в 10000 раз меньшей, чем во внеклеточной жидкости. Это обеспечивается главным образом двумя кальциевыми насосами. Один из них находится в клеточной мембране и выкачивает ионы кальция из клетки. Другой перекачивает ионы кальция в одну или более ячеистых внутриклеточных органелл, таких как саркоплазматический ретикулум в мышечных клетках или митохондрии во всех клетках. В каждом из этих случаев белок-переносчик пронизывает мембрану насквозь и функционирует как АТФ-аза, имеющая такую же способность расщеплять АТФ, как и АТФ-аза белка-переносчика ионов натрия. Различие заключается в том, что этот белок имеет высокоспецифическое место связи для кальция, а не для натрия. Первично активный транспорт ионов водорода особенно важен в двух участках тела: (1) в железах желудка; (2) в концевой части дистальных канальцев и кортикальных отделах собирательных трубочек почек. (рис.5)
В железах желудка глубоко расположенные париетальные клетки имеют самый мощный первично активный механизм для переноса ионов водорода по сравнению с любой другой частью тела. Это является основой для секреции соляной кислоты в желудке. В секреторных концах париетальных клеток желудочных желез концентрация ионов водорода повышается в миллион раз, затем они выделяются в желудок вместе с ионами хлора, формируя соляную кислоту. В почечных канальцах имеются специальные вставочные клетки в концевой части дистальных канальцев и в кортикальных отделах собирательных трубочек, где также происходит первично активный транспорт ионов водорода. В этом случае большое количество ионов водорода секретируется из крови в мочу с целью удаления избытка этих ионов из жидкостей организма против градиента концентрации, примерно в 900 раз. Количество энергии, необходимое для активного переноса вещества через мембрану, определяется степенью концентрации вещества во время переноса. Так, 100-кратное концентрирование требует энергии в 2 раза больше по сравнению с энергией, необходимой для увеличения концентрации вещества в 10 раз, а для 1000-кратного концентрирования энергии нужно в 3 раза больше. Другими словами, необходимая энергия пропорциональна десятичному логарифму степени концентрирования вещества и выражается следующей формулой: Энергия (кал/осм) = 1400 log(C1/C2) Для концентрирования 1 осмоля вещества в 10 раз нужно примерно 1400 калорий, а для концентрирования в 100 раз — 2800 калорий. Очевидно, что энергия, расходуемая для концентрирования веществ в клетках или для удаления веществ из клеток против градиента концентрации, должна быть огромной. Некоторые клетки, например выстилающие почечные канальцы и многие железистые клетки, только на эти цели тратят до 90% своей энергии. Вторично активный транспорт. Котранспорт глюкозы и аминокислот в клетке. Когда ионы натрия транспортируются наружу клетки с помощью первично активного транспорта, обычно возникает высокий концентрационный градиент ионов натрия: высокая концентрация снаружи и очень низкая концентрация внутри клетки. Этот градиент является хранилищем энергии, поскольку избыток натрия снаружи клеточной мембраны всегда пытается диффундировать внутрь клетки. При соответствующих условиях эта энергия диффузии натрия может перемещать через мембрану вместе с натрием другие вещества. Это явление называют котранспортом, оно представляет одну из форм вторично активного транспорта. Для транспорта вместе с ионами натрия другого вещества необходим механизм сопряжения. Это обеспечивается с помощью еще одного белка-переносчика в клеточной мембране. Переносчик в этом случае служит местом прикрепления, как для иона натрия, так и для транспортируемого в этом же направлении другого вещества. Когда они оба прикрепляются к белку, энергетический градиент иона натрия обеспечивает совместный перенос иона натрия и другого вещества внутрь клетки. При контртранспорте ионы натрия также пытаются диффундировать внутрь клетки в связи с их высоким градиентом концентрации. Однако на этот раз транспортируемое вещество находится внутри клетки и должно быть выведено наружу. Следовательно, ион натрия связывается с участком белка-переносчика на наружной стороне мембраны, в то время как транспортируемое в обратном направлении вещество связывается с выступающим внутрь клетки участком этого белка. Сразу после их связывания происходят конформационные изменения, и энергия, освобождаемая при движении иона натрия внутрь, обеспечивает движение другого вещества наружу. (Рис.6)
Котранспорт глюкозы и аминокислот с ионами натрия. Глюкоза и многие аминокислоты транспортируются внутрь большого числа клеток противзначительного градиента концентрации; осуществляется это исключительно механизмом котранспорта. Видно, что транспортный белок-переносчик имеет 2 участка связывания на его наружной стороне, 1 — для натрия и 1 — для глюкозы. Концентрация ионов натрия очень высокая снаружи клетки и очень низкая внутри, что обеспечивает энергию для транспорта. Транспортный белок обладает специфическим свойством: его кон формационное изменение не позволяет натрию двигаться внутрь клетки до тех пор, пока не присоединится молекула глюкозы. Когда прикрепляются оба вещества, автоматически происходит конформационное изменение белка-переносчика, в результате натрий и глюкоза одновременно транспортируются внутрь — это механизм котранспорта натрия и глюкозы. Котранспорт натрия и аминокислот происходит так же, как для глюкозы, но используются другие транспортные белки. Установлены 5 белков для транспорта аминокислоту каждый из которых способен транспортировать одну группу аминокислот со специфическими молекулярными характеристиками. Натриевый котранспорт глюкозы и аминокислот особенно характерен для эпителиальных клеток пищеварительного тракта и почечных канальцев и обеспечивает всасывание этих веществ в кровь, что будет обсуждаться в следующих главах. Другие важные механизмы котранспорта, обнаруженные в некоторых клетках, включают котранспорт ионов хлора, йода, железа и мочевой кислоты. Контртранспорт кальция и ионов водорода. Активный транспорт в тканях. Два особенно важных механизма контртранспорта (перенос в направлении, противоположном движению первичного иона) представлены натрий-кальциевым контртранспортом и натрий-водородным контртранспортом. Натрий-кальциевый контртранспорт осуществляется практически во всех клеточных мембранах, причем ионы натрия движутся внутрь, а ионы кальция — наружу, и оба иона связываются с одним транспортным белком, функционирующим в режиме контртранспорта. Этот механизм является дополнительным к первично активному транспорту кальция, который происходит в некоторых клетках. В ряде тканей осуществляется натрий-водородный контртранспорт. Наиболее важным примером являются проксимальные канальцы почек, где ионы натрия движутся из просвета канальца внутрь канальцевой клетки, тогда как протоны контртранспортом перемещаются в просвет канальца. В сравнении с первично активным транспортом протонов, происходящим в более дистальных почечных канальцах, роль контртранспортного механизма в концентрировании ионов водорода не столь значительна. Однако с его помощью может транспортироваться чрезвычайно большое число ионов водорода, что делает этот механизм ключевым для регуляции содержания этих ионов в жидкостях организма. (Рис.7)
Активный транспорт в тканях.Во многих частях организма вещества должны транспортироваться не просто через клеточную мембрану, а через слой клеток. Транспорт этого вида осуществляется через: (1) кишечный эпителий; (2) эпителий почечных канальцев; (3) эпителий экзокринных желез; (4) эпителий желчного пузыря; (5) мембрану хориоидного сплетения мозга и другие мембраны. Основными механизмами транспорта через клеточный слой являются: (1) активный транспорт через клеточную мембрану на одной стороне транспортирующих клеток; (2) простая или облегченная диффузия через мембрану на противоположной стороне этих клеток. На рисунке показан механизм транспорта ионов натрия через эпителиальный слой в кишечнике, желчном пузыре и почечных канальцах. На рисунке видно, что на стороне, обращенной в просвет, эпителиальные клетки плотно контактируют друг с другом посредством соединений, называемых легкими прикосновениями. Щеточная каемка на обращенной в просвет поверхности проницаема для ионов натрия и воды. В связи с этим натрий и вода легко диффундируют из просвета внутрь клетки. Затем ионы натрия активно транспортируются через базальную и латеральную клеточные мембраны во внеклеточную жидкость окружающей соединительной ткани и в кровеносные сосуды. В результате создается высокий градиент концентрации натрия через эти мембраны, что вызывает также осмос воды. Таким образом, активный перенос ионов натрия через базолатеральную мембрану эпителиальных клеток является причиной транспорта не только ионов натрия, но и воды. Эти механизмы обеспечивают поступление питательных веществ, ионов и других веществ из кишечника в кровь; таким же путем эти вещества реабсорбируются из клубочкового фильтрата в почечных канальцах.
Натрий-калиевый насос. К веществам, которые транспортируются посредством первично активного транспорта, относят натрий, калий, кальций, водород, хлор и некоторые другие ионы. Механизм активного транспорта лучше всего изучен для натрий-калиевого насоса (Na+/K+-нaсоса) — транспортного процесса, который выкачивает ионы натрия через мембрану клетки наружу и в то же время закачивает в клетку ионы калия. Этот насос отвечает за поддержание различной концентрации ионов натрия и калия по обе стороны мембраны, а также за наличие отрицательного электрического потенциала внутри клеток. (В главе 5 будет показано, что он является также основой процесса передачи импульсов в нервной системе.)
Белок-переносчик представлен комплексом из двух раздельных глобулярных белков: более крупного, называемого альфа-субъединицей, с молекулярной массой около 100000, и меньшего, называемого бета-субъединицей, с молекулярной массой около 55000. Хотя функция меньшего белка неизвестна (за исключением того, что он, возможно, закрепляет белковый комплекс в липидной мембране), крупный белок имеет три специфических характеристики, важные для функционирования насоса. 1. На выступающей внутрь клетки части белка имеются три рецепторных участка для связывания ионов натрия. 2. На наружной части белка располагаются два рецепторных участка для связывания ионов калия. 3. Внутренняя часть белка, расположенная вблизи участков связывания ионов натрия, обладает АТФ-азной активностью. Рассмотрим работу насоса. Когда 2 иона калия связываются с белком-переносчиком снаружи и 3 иона натрия связываются с ним внутри, активируется АТФ-азная функция белка. Это ведет к расщеплению 1 молекулы АТФ до АДФ с выделением энергии высокоэнергетической фосфатной связи. Полагают, что эта освобожденная энергия вызывает химическое и конформационное изменение молекулы белка-переносчика, в результате 3 иона натрия перемещаются наружу, а 2 иона калия — внутрь клетки. Как и другие ферменты, Na-K+-ATФ-aзa может работать и в обратном направлении. При экспериментальном увеличении электрохимических градиентов для Na+ и К+ до таких значений, что накопленная в них энергия станет выше химической энергии гидролиза АТФ, эти ионы будут двигаться по своим градиентам концентрации, а Na+/K+-Hacoc будет синтезировать АТФ из АДФ и фосфата. Следовательно, фосфорилированная форма Nа+/К+ -насоса может быть или донором фосфатов для синтеза АТФ из АДФ, или использовать энергию для изменения своей конформации и качать натрий из клетки, а калий — в клетку. Относительные концентрации АТФ, АДФ и фосфатов, как и электрохимические градиенты для натрия и калия, определяют направление ферментативной реакции. Для некоторых клеток, например, электрически активных нервных клеток, от 60 до 70% всей потребляемой клеткой энергии тратится на перемещение натрия наружу и калия внутрь. Роль Na-K-насоса: Активный транспорт ионов кальция и водорода в клетке. Одной из многочисленных важных функций Na+/K+-Hacoca является регуляция объема каждой клетки. Без функционирования этого насоса большинство клеток организма будут набухать, пока не лопнут. Механизм регуляции объема следующий: внутри клетки много белков и других органических молекул, которые не могут покинуть клетку. Большинство из них отрицательно заряжены и потому связывают большое число ионов калия, натрия и других положительно заряженных ионов. Все эти молекулы и ионы вызывают осмос воды в клетку. Без регуляции осмоса клетка будет неограниченно разбухать вплоть до разрыва мембраны. В норме механизмом для предупреждения этого является Na+/K+-Hacoc. Вспомним, что в результате работы насоса 3 иона натрия выводятся наружу, а 2 иона калия закачиваются внутрь. Кроме того, мембрана гораздо менее проницаема для ионов натрия, чем для калия, поэтому ионы натрия, оказавшись снаружи, в основном там и остаются. Следовательно, присутствует общая потеря ионов клеткой, что, в свою очередь, инициирует осмос воды из клетки. Когда клетка начинает разбухать, это автоматически активирует Na+/K+-Hacoc, обеспечивая удаление из клетки еще большего числа ионов вместе с водой. Таким образом, Na+/K+-Hacoc осуществляет непрерывную регуляцию объема клетки, поддерживая его в нормальных пределах. Электрогенная природа натрий-калиевого насоса. Как известно, Na+/K+-Hacoc выкачивает 3 иона натрия наружу на каждые 2 иона калия, входящие внутрь. Это означает, что 1 положительный заряд выводится наружу при каждом цикле работы насоса. Создается избыток положительных зарядов на поверхности клетки и дефицит положительных ионов внутри клетки, т.е. внутренняя часть клетки заряжается отрицательно. В связи с этим Na+/K+-Hacoc называют электрогенным, поскольку он создает трансмембранную разность потенциалов, а наличие электрического потенциала является основой для передачи сигналов в нервных и мышечных волокнах. Первично активный транспорт ионов кальция. Другим важным механизмом первично активного транспорта является кальциевый насос. В норме ионы кальция во внутриклеточном цитозоле практически всех клеток содержатся в чрезвычайно низкой концентрации — примерно в 10000 раз меньшей, чем во внеклеточной жидкости. Это обеспечивается главным образом двумя кальциевыми насосами. Один из них находится в клеточной мембране и выкачивает ионы кальция из клетки. Другой перекачивает ионы кальция в одну или более ячеистых внутриклеточных органелл, таких как саркоплазматический ретикулум в мышечных клетках или митохондрии во всех клетках. В каждом из этих случаев белок-переносчик пронизывает мембрану насквозь и функционирует как АТФ-аза, имеющая такую же способность расщеплять АТФ, как и АТФ-аза белка-переносчика ионов натрия. Различие заключается в том, что этот белок имеет высокоспецифическое место связи для кальция, а не для натрия. Первично активный транспорт ионов водорода особенно важен в двух участках тела: (1) в железах желудка; (2) в концевой части дистальных канальцев и кортикальных отделах собирательных трубочек почек. (рис.5)
В железах желудка глубоко расположенные париетальные клетки имеют самый мощный первично активный механизм для переноса ионов водорода по сравнению с любой другой частью тела. Это является основой для секреции соляной кислоты в желудке. В секреторных концах париетальных клеток желудочных желез концентрация ионов водорода повышается в миллион раз, затем они выделяются в желудок вместе с ионами хлора, формируя соляную кислоту. В почечных канальцах имеются специальные вставочные клетки в концевой части дистальных канальцев и в кортикальных отделах собирательных трубочек, где также происходит первично активный транспорт ионов водорода. В этом случае большое количество ионов водорода секретируется из крови в мочу с целью удаления избытка этих ионов из жидкостей организма против градиента концентрации, примерно в 900 раз. Количество энергии, необходимое для активного переноса вещества через мембрану, определяется степенью концентрации вещества во время переноса. Так, 100-кратное концентрирование требует энергии в 2 раза больше по сравнению с энергией, необходимой для увеличения концентрации вещества в 10 раз, а для 1000-кратного концентрирования энергии нужно в 3 раза больше. Другими словами, необходимая энергия пропорциональна десятичному логарифму степени концентрирования вещества и выражается следующей формулой: Энергия (кал/осм) = 1400 log(C1/C2) Для концентрирования 1 осмоля вещества в 10 раз нужно примерно 1400 калорий, а для концентрирования в 100 раз — 2800 калорий. Очевидно, что энергия, расходуемая для концентрирования веществ в клетках или для удаления веществ из клеток против градиента концентрации, должна быть огромной. Некоторые клетки, например выстилающие почечные канальцы и многие железистые клетки, только на эти цели тратят до 90% своей энергии. Вторично активный транспорт. Котранспорт глюкозы и аминокислот в клетке. Когда ионы натрия транспортируются наружу клетки с помощью первично активного транспорта, обычно возникает высокий концентрационный градиент ионов натрия: высокая концентрация снаружи и очень низкая концентрация внутри клетки. Этот градиент является хранилищем энергии, поскольку избыток натрия снаружи клеточной мембраны всегда пытается диффундировать внутрь клетки. При соответствующих условиях эта энергия диффузии натрия может перемещать через мембрану вместе с натрием другие вещества. Это явление называют котранспортом, оно представляет одну из форм вторично активного транспорта. Для транспорта вместе с ионами натрия другого вещества необходим механизм сопряжения. Это обеспечивается с помощью еще одного белка-переносчика в клеточной мембране. Переносчик в этом случае служит местом прикрепления, как для иона натрия, так и для транспортируемого в этом же направлении другого вещества. Когда они оба прикрепляются к белку, энергетический градиент иона натрия обеспечивает совместный перенос иона натрия и другого вещества внутрь клетки. При контртранспорте ионы натрия также пытаются диффундировать внутрь клетки в связи с их высоким градиентом концентрации. Однако на этот раз транспортируемое вещество находится внутри клетки и должно быть выведено наружу. Следовательно, ион натрия связывается с участком белка-переносчика на наружной стороне мембраны, в то время как транспортируемое в обратном направлении вещество связывается с выступающим внутрь клетки участком этого белка. Сразу после их связывания происходят конформационные изменения, и энергия, освобождаемая при движении иона натрия внутрь, обеспечивает движение другого вещества наружу. (Рис.6)
Котранспорт глюкозы и аминокислот с ионами натрия. Глюкоза и многие аминокислоты транспортируются внутрь большого числа клеток противзначительного градиента концентрации; осуществляется это исключительно механизмом котранспорта. Видно, что транспортный белок-переносчик имеет 2 участка связывания на его наружной стороне, 1 — для натрия и 1 — для глюкозы. Концентрация ионов натрия очень высокая снаружи клетки и очень низкая внутри, что обеспечивает энергию для транспорта. Транспортный белок обладает специфическим свойством: его кон формационное изменение не позволяет натрию двигаться внутрь клетки до тех пор, пока не присоединится молекула глюкозы. Когда прикрепляются оба вещества, автоматически происходит конформационное изменение белка-переносчика, в результате натрий и глюкоза одновременно транспортируются внутрь — это механизм котранспорта натрия и глюкозы. Котранспорт натрия и аминокислот происходит так же, как для глюкозы, но используются другие транспортные белки. Установлены 5 белков для транспорта аминокислоту каждый из которых способен транспортировать одну группу аминокислот со специфическими молекулярными характеристиками. Натриевый котранспорт глюкозы и аминокислот особенно характерен для эпителиальных клеток пищеварительного тракта и почечных канальцев и обеспечивает всасывание этих веществ в кровь, что будет обсуждаться в следующих главах. Другие важные механизмы котранспорта, обнаруженные в некоторых клетках, включают котранспорт ионов хлора, йода, железа и мочевой кислоты. Контртранспорт кальция и ионов водорода. Активный транспорт в тканях. Два особенно важных механизма контртранспорта (перенос в направлении, противоположном движению первичного иона) представлены натрий-кальциевым контртранспортом и натрий-водородным контртранспортом. Натрий-кальциевый контртранспорт осуществляется практически во всех клеточных мембранах, причем ионы натрия движутся внутрь, а ионы кальция — наружу, и оба иона связываются с одним транспортным белком, функционирующим в режиме контртранспорта. Этот механизм является дополнительным к первично активному транспорту кальция, который происходит в некоторых клетках. В ряде тканей осуществляется натрий-водородный контртранспорт. Наиболее важным примером являются проксимальные канальцы почек, где ионы натрия движутся из просвета канальца внутрь канальцевой клетки, тогда как протоны контртранспортом перемещаются в просвет канальца. В сравнении с первично активным транспортом протонов, происходящим в более дистальных почечных канальцах, роль контртранспортного механизма в концентрировании ионов водорода не столь значительна. Однако с его помощью может транспортироваться чрезвычайно большое число ионов водорода, что делает этот механизм ключевым для регуляции содержания этих ионов в жидкостях организма. (Рис.7)
Активный транспорт в тканях.Во многих частях организма вещества должны транспортироваться не просто через клеточную мембрану, а через слой клеток. Транспорт этого вида осуществляется через: (1) кишечный эпителий; (2) эпителий почечных канальцев; (3) эпителий экзокринных желез; (4) эпителий желчного пузыря; (5) мембрану хориоидного сплетения мозга и другие мембраны. Основными механизмами транспорта через клеточный слой являются: (1) активный транспорт через клеточную мембрану на одной стороне транспортирующих клеток; (2) простая или облегченная диффузия через мембрану на противоположной стороне этих клеток. На рисунке показан механизм транспорта ионов натрия через эпителиальный слой в кишечнике, желчном пузыре и почечных канальцах. На рисунке видно, что на стороне, обращенной в просвет, эпителиальные клетки плотно контактируют друг с другом посредством соединений, называемых легкими прикосновениями. Щеточная каемка на обращенной в просвет поверхности проницаема для ионов натрия и воды. В связи с этим натрий и вода легко диффундируют из просвета внутрь клетки. Затем ионы натрия активно транспортируются через базальную и латеральную клеточные мембраны во внеклеточную жидкость окружающей соединительной ткани и в кровеносные сосуды. В результате создается высокий градиент концентрации натрия через эти мембраны, что вызывает также осмос воды. Таким образом, активный перенос ионов натрия через базолатеральную мембрану эпителиальных клеток является причиной транспорта не только ионов натрия, но и воды. Эти механизмы обеспечивают поступление питательных веществ, ионов и других веществ из кишечника в кровь; таким же путем эти вещества реабсорбируются из клубочкового фильтрата в почечных канальцах.
Читайте также:
- В чем отличия обеспечения информационной безопасности в российской федерации от других стран
- Уголовное право как наука кратко
- Как зарегистрировать ребенка в рэш через госуслуги
- Что после смерти деда досталось отцу ивана сергеевича тургенева по наследству
- Обязан ли водитель включить левый указатель поворота при движении по направлению главной дороги