
Какие структуры мозга играют решающую роль в обеспечении мотивационного состояния
Обновлено: 02.07.2024
2. СТРУКТУРЫ МОЗГА, РЕАЛИЗУЮЩИЕ ПОДКРЕПЛЯЮЩУЮ, ПЕРЕКЛЮЧАЮЩУЮ, КОМПЕНСАТОРНО-ЗАМЕЩАЮЩУЮ И КОММУНИКАТИВНУЮ ФУНКЦИИ ЭМОЦИЙ
Результаты нейрофизиологических экспериментов показывают, что потребности, мотивации и эмоции имеют различный морфологический субстрат. Так, при стимуляции зон самораздражения латерального гипоталамуса градуально нарастающим электрическим током поведенческие реакции крысы всегда имеют одну и ту же последовательность. Слабое раздражение вызывает генерализованное поисковое поведение без обращения к находящимся в камере целевым объектам пище, воде, педали для самораздражения. При усилении интенсивности тока животное начинает есть, пить, грызть и т.п. По мере дальнейшего усиления тока возникает реакция самораздражения с сопутствующими мотивационными эффектами, а затем только самораздражение. Введением определённых фармакологических веществ в зоны самораздражения можно избирательно повлиять на мотивационные и эмоциональные компоненты поведения, например ослабить стремление животного к пище, не изменив объём её поглощения в случае контакта [Ikemoto, Panksepp, 1996].
Как показали эксперименты Н.Г. Михайловой и М.И. Зайченко в лаборатории П.В. Симонова, нейроны этих двух типов по-разному участвуют в реализации классических и инструментальных оборонительных условных рефлексов у крыс, когда условный сигнал (свет) сочетается с эмоционально негативным внутримозговым раздражением дорзолатеральной покрышки. На рис. 7.1 представлены изменения импульсной активности мотивационных (наверху) и позитивно подкрепляющих (внизу) нейронов латерального гипоталамуса при осуществлении условных оборонительных рефлексов: классического, избавления и избегания. Столбики последовательно показывают частоту разрядов за 5 с до условного стимула, при действии света, при действии света совместно с током и после выключения стимулов. Как следует из рисунка, осуществление классического условного рефлекса и реакции избавления сопровождается усилением активности мотивационных нейронов и подавлением импульсации подкрепляющих. Только хорошо выработанная реакция избегания, при которой крыса перестаёт получать наказание электрическим током, ведёт к усилению активности позитивно подкрепляющих нейронов. Эти данные позволяют ответить на давно обсуждаемый в литературе вопрос: что именно служит подкреплением инструментальных оборонительных рефлексов? В случае классических рефлексов и реакций избавления подкреплением служит эмоционально негативное состояние страха. При успешном осуществлении реакции избегания в процесс вовлекается механизм положительных эмоций.

Рис.7.1 . Изменения импульсной активности нейронов латерального гипоталамуса крыс при осуществлении оборонительных условных рефлексов классического (1) избавления (2) и избегания (3). По вертикали средняя частота разрядов в процентах к фону. I мотивационная. II позитивно подкрепляющая группа нейронов. Столбики последовательно представляют частоту разрядов за 5 с до включения стимулов, при действии условного света, при действии света и тока, после выключения стимулов. Одна звёздочка р меньше 0,05; две звёздочки р меньше 0,01
Оборонительные реакции нападения или бегства приурочены к различным структурам гипоталамуса. Стимуляция пунктов медиального гипоталамуса вызывающая оборонительные реакции кролика, ведёт к возрастанию количества пар скоррелированно работающих нейронов зрительной и сенсомоторной коры, причём опережение разрядами нейронов последней разрядов нейронов зрительной коры достигает 120 мс. Этот факт можно рассматривать как своеобразную подготовку зрительной области к восприятию значимых для оборонительного поведения сигналов по механизму обратных связей Э.А. Асратяна или реинтеринга Дж.Эделмена [Павлова и др., 1995].
Вовлечение эмоционально-позитивных структур в реализацию реакций избегания (но не избавления!) позволяет понять, почему сам факт активного сопротивления неблагоприятно сложившейся ситуации (например, возможность для фиксированной на спине крысы кусать палочку) достоверно ослабляет объективно регистрируемые проявления стресса.
Ранее уже говорилось о том, что влияние эмоций на поведение определяется отношением животного к своему эмоциональному состоянию, причём подчинено принципу максимизации положительных эмоций и минимизации отрицательных. Этот принцип реализуется влиянием мотивационно-эмоциональных структур гипоталамуса на информационные (когнитивные) и организующие движение отделы новой коры, о чём свидетельствует анализ пространственной синхронизации электрической активности мозговых структур при самостимуляции крыс слабым постоянным током [Pavlygina et al., 1976].
На рис 7.2 отображён процент случаев регистрации значимой когерентности (Р ПФК положительные эмоции; ПФК > ЛФК отрицательные эмоции. При подаче субъекту неосознаваемых (невербализуемых) стимулов правое полушарие более чувствительно к эмоционально отрицательным, а левое к эмоционально положительным.
Исключительно велика коммуникативная функция эмоций порождение и восприятие выражения эмоций в мимике, голосе, физических характеристиках речи. По мнению ряда исследователей, около 90% эмоционального общения происходит на неречевом (несемантическом) уровне. При восприятии речи независимо от её содержания мы можем судить об эмоциональном состоянии партнёра (например, об аффектах тоски или тревоги) по таким показателям, как средняя длительность чистой речи, продолжительность пауз, отношение времени пауз к общему времени высказывания и скорость артикуляции [Фролов и др., 1994]. Подчеркнём, что оценка эмоционального состояния партнёра по его мимике имеет собственный мозговой механизм, отличный от механизма идентификации партнёра. Так двухстороннее повреждение височной коры нарушает узнавание знакомого лица, а одностороннее повреждение правой височной области достаточно для нарушения распознавания эмоциональной экспрессии. Двухстороннее повреждение миндалин, напротив, препятствует распознаванию мимики страха, не влияя на идентификацию знакомых и незнакомых лиц [Adolphs et al., 1995], а также интонационных признаков страха и гнева.
В процессах порождения и восприятия эмоциональной мимики мы вновь встречаемся с функциональной асимметрией мозга. Эмоциональное состояние субъекта преимущественно отражается на мимике левой половины лица, что свидетельствует о преобладающей активности правого полушария. Аналогичные данные получены и в опытах на макаках, что указывает на общность обнаруженной закономерности для всех приматов. Естественно, что эмоции распознаются лучше на фотографиях лиц, составленных из двух левых половин изображения. Правое полушарие (его центральная височная область) преобладает и при восприятии эмоциональной мимики. Правда, тщательно выполненное исследование обнаруживает достаточно сложную и динамичную картину взаимодействия полушарий при опознании эмоциональной лицевой экспрессии. При правильном опознании выражений радости, печали или эмоционально нейтральных лиц последовательно активируются фронтальная кора правого полушария, а затем фронтальная кора левого. При ошибочных опознаниях активация левого полушария опережает активацию правого [Михайлова и др., 1996].
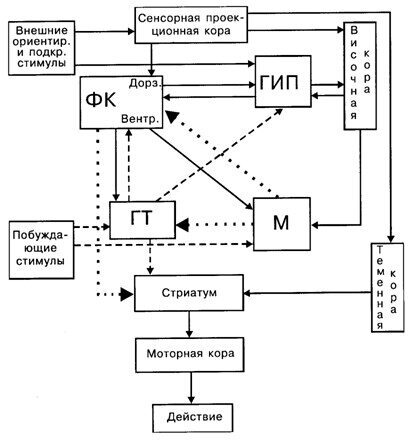
Рис. 7.5. Схема взаимодействия мозговых структур в процессе организации поведенческого акта: ФК– фронтальная кора; ГИП– гиппокомп; М– миндалина; ГТ– гипоталамус; сплошные линии информационная афферентация, пунктирные мотивационные влияния, точечные эмоционально окрашенная афферентация
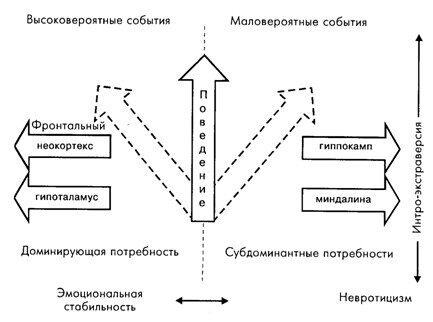
Рис. 7.6. Гипотетическая схема зависимости параметров интро-экстраверсии и эмоциональной стабильности невротицизма от индивидуальных особенностей взаимодействия мозговых структур

В 1937 г нейроанатом Джеймс Папец обратил внимание на существование многочисленных связей между структурами мозга, расположенными в виде кольца (лимба) в области соединения ствола и полушарий, и предложил объединить их в лимбическую систему. Кольцо лимбической системы образуют мамиллярные тела, гиппокамп, миндалины и свод, проходящий дугой от гиппокампа к мамиллярным телам и перегородке. В лимбическую систему принято также включать соседние области древней и старой коры мозга: прилегающие к гиппокампу грушевидные доли и поясные извилины – они занимают медиальную поверхность больших полушарий непосредственно над мозолистым телом и, как пояс, огибают таламус. Лимбические структуры связаны хорошо развитыми проводящими путями с обонятельным мозгом и гипоталамусом, кортикальными центрами познания и эмоций (рис. 13.3). Изучение функций лимбической системы позволило обнаружить различные нейромедиаторы и рецепторы у нейронов, входящих в состав лимбической системы.

Американский физиолог Пол Мак-Лин предложил модель иерархического деления мозга на три отдела в соответствии с этапами его эволюционного развития (рис. 13.4): 1) древний мозг рептилий (ствол, промежуточный мозг и базальные ганглии); 2) старый мозг млекопитающих (структуры лимбической системы) и 3) новый мозг млекопитающих (кора больших полушарий).
Функция древнего мозга заключается в контроле врождённых поведенческих актов; такой мозг недостаточно пластичен и обеспечивает выживание только при постоянных условиях среды. Древний мозг формирует эмоции, увеличивает объём памяти и даёт возможность возникновения простых форм поведения. Новый мозг прибавляет возможность произвольного управления эмоциями, прогнозирования поведения и т.п.
В случае если одновременно возникают несколько мотиваций, необходимо выбрать одну из них — наиболее насущную. Если речь идет о поведении простейших существ, у которых набор потребностей невелик (пищевая, питьевая, половая и самосохранение), то такой конфликт между мотивациями можно решить путем врожденной иерархии потребностей (потребность в самосохранении всегда преобладает над остальными) или по принципу фактора силы — возникает то поведение, которое запускается более сильным раздражителем. Однако для человека с его чрезвычайно разнообразными и сложными потребностями такой способ выбора поведения невозможен. Следовательно, необходим другой механизм разрешения конфликта мотиваций, и этим механизмом являются эмоции. При выборе поведения высшие живые существа руководствуются универсальным принципом:
— чем важнее потребность, тем более положительную эмоцию дает ее удовлетворение (или тем больше устраняет отрицательную эмоцию);
— живые существа выбирают такое поведение, которое вызовет наиболее сильную положительную эмоцию или сведет к минимуму отрицательную эмоцию.
Основные структуры мозга, отвечающие за эмоции и мотивации, сосредоточены в лимбической системе. Её функции чрезвычайно сложны и разнообразны, но тем не менее все они являются частью эмоционально-мотивационного поведения или, по крайней мере, тесно связаны с ним.

На рис. 18.8 приведена схема распределенной системы эмоционально-мотивационного поведения (распределенные системы — это системы, объединяющие нервные центры на разных уровнях ЦНС для выполнения сложной функции). Эмоционально-мотивационное поведение возникает следующим образом:
- потребность воспринимается ЦНС, превращаясь в мотивацию;
- мотивация вызывает активацию эмоциогенных структур, то есть эмоциогенное возбуждение;
- эмоциогенное возбуждение вызывает цельную эмоцию, проявляющуюся эмоциональным выражением и эмоциональным переживанием;
- определяя значимость побуждения, эмоция обеспечивает выбор ведущей мотивации;
- ведущая мотивация запускает поведение.
Из всего этого следует, что в ЦНС должны существовать структуры:
- воспринимающие потребности, превращая их в мотивации;
- отвечающие за эмоциогенное возбуждение;
- отвечающие за эмоциональное выражение;
- отвечающие за эмоциональное переживание.
В гипоталамусе происходит восприятие потребности и превращение ее в мотивацию – активацию мотивационных центров гипоталамуса. Поражения этих центров приводят к полному распаду мотивационного поведения или его неконтролируемому усилению (анорексия, булимия, полидипсия).
В эмоциогенных зонах лимбической системы возникает эмоциогенное возбуждение – пусковой механизм развития эмоции. Соответственно, при поражениях этих структур возникают эмоциональные расстройства, например:
- депрессия – постоянно подавленное настроение, преобладание тяжелых отрицательных эмоций;
- мания – постоянно повышенное настроение, эйфория, бурные и неконтролируемые положительные эмоции.
В среднем мозге заложены программы эмоционального выражения. При перерыве путей к среднему мозгу, отвечающих за запуск этих программ, с одной стороны, эмоции не сопровождаются характерным выражением (например, мимикой), с другой — эмоциональное выражение может возникать без соответствующей эмоции (рефлекторно, под влиянием действующих на средний мозг раздражителей), например:
Роль миндалин в мотивации
Если гипоталамус является важнейшей мотивирующей структурой мозга, активирующейся при изменении постоянства внутренней среды, то на внешние стимулы, вызывающие мотивацию или же изменяющие уже мотивированное поведение, раньше гипоталамуса откликаются нейроны миндалин мозга, которые относятся к лимбической системе и находятся в височных долях больших полушарий.

Есть два важных источника афферентной информации для миндалин: сенсорные ядра таламуса и кора, преимущественно вторичные слуховые области и полимодальные ассоциативные поля (рис. 13.8). При действии внешних стимулов сенсорная информация раньше поступает от таламуса и вызывает примитивную реакцию, на фоне которой приходит информация, уже переработанная в коре. Оба афферентных потока направляются к базолатеральным ядрам миндалин, где пришедшие сигналы перерабатываются и передаются нейронам центрального ядра миндалин. От клеток центрального ядра начинаются эфферентные пути миндалин, которые направляются к латеральному гипоталамусу и стволу мозга. При возникновении страха активация миндалин приводит к последующему быстрому повышению частоты сокращений сердца и артериального давления, учащению дыхания, выделению гормонов и другим проявлениям симпатоадреналовой реакции и стресса.
Помимо этого существует эфферентный путь от миндалин к коре, в особенности к орбитофронтальной области и поясной извилине, этот путь важен для осознания переживаемых ощущений. Ядра миндалин имеют реципрокные соединения с гиппокампом и гипоталамусом.
У кроликов можно выработать условный рефлекс на нейтральный звуковой стимул, подкрепляя его болевым действием электрического тока. Болевое раздражение всегда сопровождается повышением частоты сокращений сердца, артериального давления, учащением дыхания и т.п. Когда образуется условный рефлекс, такая же реакция наблюдается и на прежде нейтральный звуковой раздражитель, который начинает вызывать у животного состояние страха. Если удалить (блокировать) миндалины – реакция страха исчезнет. Т.е. не только врождённые, но и приобретённые мотивации страха связаны с обязательным участием миндалин.
Роль миндалин не сводится только к мотивации страха. Хотя при их повреждении не исчезают совсем мотивации голода или жажды, но в пищевом и питьевом поведении наступают заметные перемены. Подопытные животные перестают отличать предпочитаемую раньше вкусную пищу от невкусной и постоянно берут в рот даже несъедобные предметы. У них пропадает врождённая или выработанная до операции разборчивость в питье – подопытные крысы начинают пить растворы с неприятным для нормальных животных запахом. У них изменяется половое поведение: животные становятся гиперсексуальными и готовы спариваться даже с представителями другого вида. Т.е. значение миндалин становится особенно заметным в тех случаях, когда для формирования мотивированного поведения одновременно требуется учесть существование нескольких внешних факторов (главная функция миндалины – выделение доминирующей потребности, подлежащей первоочередному удовлетворению).
Резюме
Изменения гомеостатических параметров приводят в действие нервные и эндокринные механизмы их регуляции, направленные на восстановление заданного значения параметров. Одновременно с этим или заблаговременно формируется специфическое поведение, направленное на достижение той же цели. Главной мотивационной структурой мозга является гипоталамус, нейроны которого активируются притоком специфических афферентных импульсов и прямым влиянием гуморальных факторов. В мотивациях, связанных с действием внешних факторов важную роль играют миндалины мозга. Конкретные формы мотивированного поведения и определения мотивационной доминанты определяются сложными разновидностями взаимодействия между подкорковыми структурами (лимбической системой) и корой мозга, причём важную роль играет прежний поведенческий опыт и следы памяти.

Важным анатомическим фактом, является неравномерность распределения отдельных слоев коры в топографически различных участках коры головного мозга.
Факты показывают, что над каждой первичной областью мозговой коры, в которой преобладают низшие (афферентные или эфферентные) слои коры, надстраиваются вторичные области, в которых преобладают верхние (проекционно-ассоциационные) слои, играющие, существенную роль в функциональной организации работы отдельных анализаторов.
Концепция структурно-функциональных блоков мозга А. Р. Лурия
А. Р. Лурия предложил структурно-функциональную модель мозга как субстрата психической деятельности. Эта модель характеризует наиболее общие закономерности работы мозга как единого целого и позволяет объяснить его интегративную функцию (Е. Д. Хомская).
Согласно этой модели, весь мозг можно разделить на три структурно-функциональных блока:
а) Энергетический блок,
б) Блок приема, переработки и хранения экстероцептивной информации,
в) Блок программирования, регуляции и контроля сложных форм деятельности.
Любая ВПФ осуществляется при обязательном участии всех трех блоков. Каждый блок характеризуется особенностями строения, физиологическими принципами, лежащими в основе его работы, и той ролью, которую он играет в обеспечении психических функций .
Первый функциональный блок мозга
Первый блок — это блок регуляции энергетического тонуса и бодрствования.
Было доказано (И. П. Павлов, А. Р. Лурия, М. Н. Ливанов), что для нормальной психической деятельности организм должен находиться в состоянии бодрствования (иными словами, кора больших полушарий должна находиться в состоянии тонуса, т. е. иметь определенный уровень возбуждения).
Только в условиях оптимального бодрствования человек может наилучшим образом принимать и перерабатывать информацию, вызывать в памяти нужные системы связей, программировать деятельность, осуществлять контроль над ней.
Было установлено, что аппараты, обеспечивающие и регулирующие тонус коры, находятся не в самой коре, а в лежащих ниже стволовых и корковых отделах мозга.
Таким аппаратом являются неспецифические структуры разных уровней:
- ретикулярная формация ствола мозга,
- неспецифические структуры среднего мозга,
- лимбическая система,
- область гиппокампа.
Ретикулярная формация представляет собой нервную сеть, в которую вкраплены тела нейронов с короткими аксонами.

Ретикулярная формация имеет ряд особенностей строения и функционирования, благодаря которым обеспечиваются ее основные функции:
Наличие многочисленных связей в самой ретикулярной формации, конвергенция всех нервных путей на большей части ее нейронов создают дополнительные возможности широкого и одновременного распространения волн возбуждения в первичные, вторичные и третичные зоны коры, а также другие структуры мозга.
Как известно, нервная система всегда находится в состоянии определенной активности и для любого проявления жизнедеятельности обязательно ее наличие.
Принято выделять несколько источников активности: в первую очередь, обменные процессы организма, лежащие в основе гомеостаза (белковый, углеводный и т. д.). Затем непосредственный приток информации, поступающей в организм из внешнего мира (от экстерорецепторов).
Известно, что в состоянии сенсорной депривации человек впадает в сон, из которого его может вывести лишь поступление новой информации.
Перечисленные источники активности свойственны и человеку, и животным. Но у человека помимо этого значительная часть активности обусловлена его планами, намерениями, программами. Формируясь в процессе сознательной жизни, они являются социальными по своему заказу и осуществляются при ближайшем участии сначала внешней, а потом внутренней речи.
Функциональное значение первого блока в обеспечении психических функций заключается,
- Во-первых, в регуляции процессов активации, в поддержании общего тонуса ЦНС, необходимого для любой психической деятельности (активирующая функция).
- Во-вторых, в передаче регулирующего влияния мозговой коры на нижележащие стволовые образования (модулирующая функция): За счет нисходящих волокон ретикулярной формации высшие отделы коры управляют работой нижележащих аппаратов, модулируя их работу и обеспечивая сложные формы сознательной деятельности.
При поражении ретикулярной формации снижается продуктивность всех ВПФ (в первую очередь — непроизвольного внимания и памяти), нарушается активность, сон.
В случае массивных поражений ретикулярной формации стирается грань между сном и бодрствованием, человек находится в полусонном состоянии, у него страдает ориентировка во времени и месте.
Отличительными диагностическими признаками поражения ретикулярной формации является одновременное снижение продуктивности абсолютно всех психических процессов, а также возможность частичной компенсации дефекта за счет усложнения задания.
Вовлечение произвольных процессов и специальная мотивация позволяют ненадолго повысить эффективность психических процессов.
Таким образом, первый блок мозга участвует в обеспечении психической деятельности, в первую очередь в организации внимания, памяти, эмоционального состояния и сознания в целом.
Кроме того, первый блок мозга участвует в регуляции эмоциональных (страх, боль, удовольствие, гнев) и мотивационных состояний.
Лимбические структуры мозга, входящие в этот блок, занимают центральное место в организации эмоциональных и мотивационных состояний.
В связи с этим первый блок мозга воспринимает и перерабатывает разнообразную интероцептивную информацию о состоянии внутренних органов и регулирует эти состояния.
Второй функциональный блок мозга
Второй блок — блок приема, переработки и хранения информации – расположен в наружных отделах новой коры (неокортекса) и занимает ее задние отделы, включая в свой состав аппараты затылочной, височной и теменной коры.

Структурно-анатомической особенностью этого блока мозга является шестислойное строение коры.
- первичные зоны (обеспечивающие прием и анализ поступающей извне информации),
- вторичные зоны (выполняющие функции синтеза информации от одного анализатора) и
- третичные зоны (основной задачей которых является комплексный синтез информации).
Отличительной особенностью аппаратов второго блока является модальная специфичность.
Эксперименты по регистрации активности отдельных нейронов показали, что нервные клетки первичных зон отличаются высокой модальной специфичностью и узкой специализацией.
Первое означает, что они реагирует на возбуждение только одной модальности (одного вида), например, только зрительное или только слуховое.
Второе предполагает, что эти нейроны реагируют лишь на отдельный признак раздражителя одного вида (например, только на ширину линии или угол наклона и т. п.).
Благодаря этому аппараты второго функционального блока мозга выполняют функции приема и анализа информации, поступающей от внешних рецепторов и синтеза этой информации.
Основные законы построения коры, входящей в состав второго блока мозга
А. Р. Лурия выделяет основные законы построения коры, входящей в состав второго блока мозга.
1. Закон иерархического строения корковых зон.
Согласно этому закону соотношение первичных, вторичных и третичных зон коры осуществляет все более сложный синтез информации.
Более сложно организованные зоны коры обеспечивают более сложные функции.
А. Р. Лурия подчеркивает, что соотношение первичных, вторичных и третичных зон у взрослого и у ребенка различно.
Для нормального развития вторичных зон у ребенка необходимо, чтобы были сформированы первичные, а для развития третичных — вторичные зоны.
Поэтому поражение первичных зон в раннем детстве может приводить к грубым нарушениям в развитии вторичных и, тем более, третичных зон.
У взрослого же человека, при сформированных зонах коры, третичные, наиболее организованные, управляют функцией ниже лежащих вторичных и первичных зон. Поэтому у взрослого человека взаимодействие зон коры осуществляется сверху вниз.
В данном случае поражение первичных зон не приводит к заметным нарушениям психических функций и может компенсироваться работой расположенных рядом структур.
2. Закон убывающей модальной специфичности иерархически построенных корковых зон.
Этот закон предполагает, что по мере перехода от первичных зон к третичным снижается проявление их модальной специфичности.
Первичные зоны каждой из долей мозга, входящих во второй блок мозга, обладают максимальной модальной специфичностью (благодаря громадному числу нейронов с высокодифференцированной, модально-специфической функцией).
Вторичные зоны, в которых преобладают верхние слои с ассоциативными нейронами, обладают модальной специфичностью в значительно меньшей степени.
Таким образом, этот закон описывает переход от дробного отражения частных, модально-специфических признаков к синтетическому отражению более общих схем воспринимаемого мира.
3. Закон прогрессивной латерализации функций.
Закон прогрессивной латерализации функций объясняет связь функций с определенным полушарием (по мере перехода от первичных зон к третичным зонам).
Первичные зоны обоих полушарий мозга равноценны. На уровне вторичных зон часть функций, выполняемых левым и правым полушариями, остаются одинаковыми, но часть функций левого полушария уже отличаются от функций, выполняемых правым полушарием мозга.
Функции же третичных зон левого полушария уже коренным образом отличаются от функций аналогичных зон правого полушария мозга.
При поражении аппаратов второго блока мозга нарушение функций зависит от того, какие именно зоны пострадали.
При поражении первичных зон возникает нарушение восприятия отдельных признаков воспринимаемого раздражителя одной модальности (слепое пятно, гемеанопсия, нарушение тон-шкалы, анестезия и т. д.).
При поражении вторичных зон коры наблюдается нарушение синтеза отдельных признаков воспринимаемого раздражителя в целостный образ одной модальности (агнозии, афазии).
Поражение третичных зон приводит к нарушению комплексного синтеза раздражений, поступающих от разных анализаторов, что проявляется в нарушении ориентировки в пространстве.
Причем, согласно закону прогрессивной латерализации, при поражении третичных зон правого полушария нарушается предметная ориентировка в пространстве, а при поражении аналогичных зон левого полушария — страдает символическая ориентировка в пространстве.
Третий функциональный блок мозга
Третий функциональный блок мозга — блок программирования, регуляции и контроля сложных форм деятельности.
Он связан с организацией целенаправленной, сознательной психической активности, которая включает в свою структуру цель, мотив, программу действий по достижению цели, выбор средств, контроль за выполнением действий, коррекцию полученного результата. Обеспечению этих задач и служит третий блок мозга.
Аппараты третьего функционального блока мозга расположены кпереди от центральной лобной извилины и включают в свой состав моторные, премоторные и префронтальные отделы коры лобных долей мозга.

Лобные доли отличаются очень сложным строением и большим числом двусторонних связей со многими корковыми и подкорковыми структурами.
Отличительной особенностью этого блока является:
- Проведение процессов возбуждения от третичных зонах к вторичным, затем к первичным;
- Отсутствие модально-специфических зон (состоит из аппаратов только двигательного типа);
- Наличие обширных двусторонних связей не только с нижележащими образованиями ствола мозга, но и со всеми остальными отделами коры больших полушарий.
По своей структуре и функциональной организации моторная кора относится к первичным, премоторная — к вторичным, а префронтальная — к третичным зонам коры больших полушарий.
Поэтому они выполняют функции, характерные для этих зон.

Премоторная кора обеспечивает двигательные программы, т. е. объединяет отдельные движения в единую кинетическую мелодию.
Префронтальные отделы играют решающую роль в формировании намерений, программ, в регуляции и контроле наиболее сложных форм поведения человека. Они состоят из мелкозернистых клеток с короткими аксонами и обладают мощными пучками восходящих и нисходящих связей с ретикулярной формацией.
Поэтому могут выполнять ассоциативную функцию, получая импульсы от первого блока мозга и оказывать интенсивное модулирующее влияние на образования ретикулярной формации, приводя ее активирующие импульсы в соответствие с динамическими схемами поведения, которые формируются непосредственно в префронтальной (лобной) коре.
Префронтальные отделы фактически надстроены над всеми отделами мозговой коры, выполняя функцию общей регуляции поведения.
При поражении третьего функционального блока мозга характер нарушения функций связан с тем, какой именно аппарат поврежден.
При поражении моторной коры затрудняется проведение возбуждения к конкретным мышцам (наблюдаются парезы и параличи отдельных групп мышц).
Поражение премоторной коры приводит к нарушению синтеза отдельных движений в единое целое (распад двигательных навыков), поражение префронтальных отделов проявляется в нарушении сознательной целенаправленной деятельности.
Анализ особенностей строения и функционирования трех функциональных блоков мозга позволяет предположить, что каждая форма сознательной деятельности всегда является сложной функциональной системой и осуществляется, опираясь на совместную работу всех трех блоков мозга, каждый из которых вносит свой вклад в обеспечение всего психического процесса в целом.
Выделение этих блоков достаточно условно.
Аппараты первого блока мозга обеспечивают необходимый уровень активации остальных блоков, второй блок отвечает за прием, анализ и переработку информации, поступающей из внешней среды, от экстерорецепторов.
С одной стороны, эта информация составляет основу для интегративной деятельности третьего блока, а с другой — является одним из источников активности первого блока.
Роль третьего блока проявляется в его интегрирующей, регулирующей, модулирующей функции. Обеспечивая важный этап в переработке информации, аппараты третьего блока оказывают модулирующее влияние и на первый и на второй блоки, что приводит как к изменению уровня активации коры, с одной стороны, так и к изменению порогов ощущения и восприятия — с другой.
Анализируя взаимодействие блоков мозга с точки зрения деятельности, следует отметить, что первый блок участвует в формировании мотивов любой сознательной деятельности, второй — обеспечивает операциональную сторону деятельности, а третий — отвечает за формирование целей и программ деятельности.
Нарушение работы каждого из этих блоков обязательно приводит к дезинтеграции психической деятельности в целом, но каждый раз по-разному, так как приводит к нарушению соответствующих стадий деятельности.
Свидетельство и скидка на обучение каждому участнику
Концепция структурно-функциональных блоков мозга А.Р. Лурия.
А. Р. Лурия предложил структурно-функциональную модель мозга как субстрата психической деятельности. Эта модель характеризует наиболее общие закономерности работы мозга как единого целого и позволяет объяснить его интегративную функцию (Е. Д. Хомская). Согласно этой модели, весь мозг можно разделить на три структурно-функциональных блока: а) энергетический блок, б) блок приема, переработки и хранения экстероцептивной информации, в) блок программирования, регуляции и контроля сложных форм деятельности. Любая ВПФ осуществляется при обязательном участии всех трех блоков. Каждый блок характеризуется особенностями строения, физиологическими принципами, лежащими в основе его работы, и той ролью, которую он играет в обеспечении психических функций.
Первый блок - это блок регуляции энергетического тонуса и бодрствования. Было доказано (И. П. Павлов, А. Р. Лурия, М. Н. Ливанов), что для нормальной психической деятельности организм должен находиться в состоянии бодрствования (иными словами, кора больших полушарий должна находиться в состоянии тонуса, т. е. иметь определенный уровень возбуждения). Только в условиях оптимального бодрствования человек может наилучшим образом принимать и перерабатывать информацию, вызывать в памяти нужные системы связей, программировать деятельность, осуществлять контроль над ней. Было установлено, что аппараты, обеспечивающие и регулирующие тонус коры, находятся не в самой коре, а в лежащих ниже стволовых и корковых отделах мозга. Таким аппаратом являются неспецифические структуры разных уровней - ретикулярная формация ствола мозга, неспецифические структуры среднего мозга, лимбическая система, область гиппокампа. Ретикулярная формация представляет собой нервную сеть, в которую вкраплены тела нейронов с короткими аксонами.
Ретикулярная формация имеет ряд особенностей строения и функционирования, благодаря которым обеспечиваются ее основные функции: во-первых, она состоит из восходящей и нисходящей частей. По волокнам восходящей ретикулярной формации возбуждение направляется вверх, оканчиваясь в расположенных выше образованиях (гипоталамусе, древней коре и новой коре). Волокна нисходящей ретикулярной формации имеют обратное направление: начинаясь от новой коры, они передают возбуждение к структурам среднего мозга и ствола мозга. Кроме того, нейроны ретикулярной формации работают по принципу "постепенного накопления возбуждения", т. е. возбуждение распространяется не отдельными импульсами, а градуально, постепенно меняя свой уровень и таким образом модулируя состояние всего нервного аппарата. И, наконец, к ретикулярной формации сходятся волокна (колатерали) от всех анализаторных систем, а также волокна из коры головного мозга и мозжечка. Наличие многочисленных связей в самой ретикулярной формации, конвергенция всех нервных путей на большей части ее нейронов создают дополнительные возможности широкого и одновременного распространения волн возбуждения в первичные, вторичные и третичные зоны коры, а также другие структуры мозга.
Как известно, нервная система всегда находится в состоянии определенной активности и для любого проявления жизнедеятельности обязательно ее наличие. Принято выделять несколько источников активности: в первую очередь, обменные процессы организма, лежащие в основе гомеостаза (белковый, углеводный и т. д.). Затем непосредственный приток информации, поступающей в организм из внешнего мира (от экстерорецепторов). Известно, что в состоянии сенсорной депривации человек впадает в сон, из которого его может вывести лишь поступление новой информации. Перечисленные источники активности свойственны и человеку, и животным. Но у человека помимо этого значительная часть активности обусловлена его планами, намерениями, программами. Формируясь в процессе сознательной жизни, они являются социальными по своему заказу и осуществляются при ближайшем участии сначала внешней, а потом внутренней речи.
Е. Д. Хомская подчеркивает, что функциональное значение первого блока в обеспечении психических функций заключается, во-первых, в регуляции процессов активации, в поддержании общего тонуса ЦНС, необходимого для любой психической деятельности (активирующая функция). Во-вторых, в передаче регулирующего влияния мозговой коры на нижележащие стволовые образования (модулирующая функция). За счет нисходящих волокон ретикулярной формации высшие отделы коры управляют работой нижележащих аппаратов, модулируя их работу и обеспечивая сложные формы сознательной деятельности.
При поражении ретикулярной формации снижается продуктивность всех ВПФ (в первую очередь - непроизвольного внимания и памяти), нарушается активность, сон. В случае массивных поражений стирается грань между сном и бодрствованием, человек находится в полусонном состоянии, у него страдает ориентировка во времени и месте. Отличительными диагностическими признаками поражения ретикулярной формации является одновременное снижение продуктивности абсолютно всех психических процессов, а также возможность частичной компенсации дефекта за счет усложнения задания. Вовлечение произвольных процессов и специальная мотивация позволяют ненадолго повысить эффективность психических процессов.
Таким образом, первый блок мозга участвует в обеспечении психической деятельности, в первую очередь в организации внимания, памяти, эмоционального состояния и сознания в целом. Кроме того, первый блок мозга участвует в регуляции эмоциональных (страх, боль, удовольствие, гнев) и мотивационных состояний. Лимбические структуры мозга, входящие в этот блок, занимают центральное место в организации эмоциональных и мотивационных состояний. В связи с этим первый блок мозга воспринимает и перерабатывает разнообразную интероцептивную информацию о состоянии внутренних органов и регулирует эти состояния.
Второй блок - блок приема, переработки и хранения информации расположен в наружных отделах новой коры (неокортекса) и занимает ее задние отделы, включая в свой состав аппараты затылочной, височной и теменной коры. Структурно-анатомической особенностью этого блока мозга является шестислойное строение коры. Она включает первичные зоны (обеспечивающие прием и анализ поступающей извне информации), вторичные зоны (выполняющие функции синтеза информации от одного анализатора) и третичные зоны (основной задачей которых является комплексный синтез информации).
Отличительной особенностью аппаратов второго блока является модальная специфичность. Эксперименты по регистрации активности отдельных нейронов показали, что нервные клетки первичных зон отличаются высокой модальной специфичностью и узкой специализацией. Первое означает, что они реагирует на возбуждение только одной модальности (одного вида), например, только зрительное или только слуховое. Второе предполагает, что эти нейроны реагируют лишь на отдельный признак раздражителя одного вида (например, только на ширину линии или угол наклона и т. п.). Благодаря этому аппараты второго функционального блока мозга выполняют функции приема и анализа информации, поступающей от внешних рецепторов и синтеза этой информации.
А. Р. Лурия выделяет основные законы построения коры, входящей в состав второго блока мозга. Закон иерархического строения корковых зон. Согласно этому закону соотношение первичных, вторичных и третичных зон коры осуществляет все более сложный синтез информации. Более сложно организованные зоны коры обеспечивают более сложные функции. А. Р. Лурия подчеркивает, что соотношение первичных, вторичных и третичных зон у взрослого и у ребенка различно. Для нормального развития вторичных зон у ребенка необходимо, чтобы были сформированы первичные, а для развития третичных - вторичные зоны. Поэтому поражение первичных зон в раннем детстве может приводить к грубым нарушениям в развитии вторичных и, тем более, третичных зон. У взрослого же человека, при сформированных зонах коры, третичные, наиболее организованные, управляют функцией ниже лежащих вторичных и первичных зон. Поэтому у взрослого человека взаимодействие зон коры осуществляется сверху вниз. В данном случае поражение первичных зон не приводит к заметным нарушениям психических функций и может компенсироваться работой расположенных рядом структур.
Закон убывающей модальной специфичности иерархически построенных корковых зон предполагает, что по мере перехода от первичных зон к третичным снижается проявление их модальной специфичности. Первичные зоны каждой из долей мозга, входящих во второй блок мозга, обладают максимальной модальной специфичностью (благодаря громадному числу нейронов с высокодифференцированной, модально-специфической функцией). Вторичные зоны, в которых преобладают верхние слои с ассоциативными нейронами, обладают модальной специфичностью в значительно меньшей степени. Еще меньше модальная специфичность характерна для третичных зон описываемого блока ("зоны перекрытия" корковых концов различных анализаторов). Таким образом, этот закон описывает переход от дробного отражения частных, модально-специфических признаков к синтетическому отражению более общих схем воспринимаемого мира.
Закон прогрессивной латерализации функций объясняет связь функций с определенным полушарием (по мере перехода от первичных зон к третичным зонам). Первичные зоны обоих полушарий мозга равноценны. На уровне вторичных зон часть функций, выполняемых левым и правым полушариями, остаются одинаковыми, но часть функций левого полушария уже отличаются от функций, выполняемых правым полушарием мозга. Функции же третичных зон левого полушария уже коренным образом отличаются от функций аналогичных зон правого полушария мозга.
Е. Д. Хомская указывает, что при поражении аппаратов второго блока мозга нарушение функций зависит от того, какие именно зоны пострадали. При поражении первичных зон возникает нарушение восприятия отдельных признаков воспринимаемого раздражителя одной модальности (слепое пятно, гемеанопсия, нарушение тон-шкалы, анестезия и т. д.). При поражении вторичных зон коры наблюдается нарушение синтеза отдельных признаков воспринимаемого раздражителя в целостный образ одной модальности (агнозии, афазии). Поражение третичных зон приводит к нарушению комплексного синтеза раздражений, поступающих от разных анализаторов, что проявляется в нарушении ориентировки в пространстве. Причем, согласно закону прогрессивной латерализации, при поражении третичных зон правого полушария нарушается предметная ориентировка в пространстве, а при поражении аналогичных зон левого полушария - страдает символическая ориентировка в пространстве.
Третий функциональный блок мозга - блок программирования, регуляции и контроля сложных форм деятельности. Он связан с организацией целенаправленной, сознательной психической активности, которая включает в свою структуру цель, мотив, программу действий по достижению цели, выбор средств, контроль за выполнением действий, коррекцию полученного результата. Обеспечению этих задач и служит третий блок мозга.
Аппараты третьего функционального блока мозга расположены кпереди от центральной лобной извилины и включают в свой состав моторные, премоторные и префронтальные отделы коры лобных долей мозга. Лобные доли отличаются очень сложным строением и большим числом двусторонних связей со многими корковыми и подкорковыми структурами. Отличительной особенностью этого блока является проведение процессов возбуждения от третичных зонах к вторичным, затем к первичным; отсутствие модально-специфических зон (состоит из аппаратов только двигательного типа); наличие обширных двусторонних связей не только с нижележащими образованиями ствола мозга, но и со всеми остальными отделами коры больших полушарий.
По своей структуре и функциональной организации моторная кора относится к первичным, премоторная - к вторичным, а префронтальная - к третичным зонам коры больших полушарий. Поэтому они выполняют функции, характерные для этих зон. Нейроны моторной коры передают возбуждение к мышцам, отсюда начинается большой пирамидный путь. Именно эти зоны имеют ярко выраженную соматотопическую организацию, что наглядно проиллюстрировал Пенфилд ("двигательный человечек"). Премоторная кора обеспечивает двигательные программы, т. е. объединяет отдельные движения в единую кинетическую мелодию. Префронтальные отделы играют решающую роль в формировании намерений, программ, в регуляции и контроле наиболее сложных форм поведения человека. Они состоят из мелкозернистых клеток с короткими аксонами и обладают мощными пучками восходящих и нисходящих связей с ретикулярной формацией. Поэтому могут выполнять ассоциативную функцию, получая импульсы от первого блока мозга и оказывать интенсивное модулирующее влияние на образования ретикулярной формации, приводя ее активирующие импульсы в соответствие с динамическими схемами поведения, которые формируются непосредственно в префронтальной (лобной) коре. Префронтальные отделы фактически надстроены над всеми отделами мозговой коры, выполняя функцию общей регуляции поведения.
При поражении третьего функционального блока мозга характер нарушения функций связан с тем, какой именно аппарат поврежден. При поражении моторной коры затрудняется проведение возбуждения к конкретным мышцам (наблюдаются парезы и параличи отдельных групп мышц). Поражение премоторной коры приводит к нарушению синтеза отдельных движений в единое целое (распад двигательных навыков), поражение префронтальных отделов проявляется в нарушении сознательной целенаправленной деятельности. Исследования А. Р. Лурия показали, что в этом случае целенаправленное поведение заменяется "полевым" поведением, нарушается программа выполняемой деятельности, лобный больной не может подобрать адекватные средства деятельности, отсутствует контроль за ее выполнением и коррекция ошибок. При этом наблюдается персеверация (повторяемость) и стереотипизация движений.
Анализ особенностей строения и функционирования трех функциональных блоков мозга позволяет предположить, что каждая форма сознательной деятельности всегда является сложной функциональной системой и осуществляется, опираясь на совместную работу всех трех блоков мозга, каждый из которых вносит свой вклад в обеспечение всего психического процесса в целом. Выделение этих блоков достаточно условно
Аппараты первого блока мозга обеспечивают необходимый уровень активации остальных блоков, второй блок отвечает за прием, анализ и переработку информации, поступающей из внешней среды, от экстерорецепторов. С одной стороны, эта информация составляет основу для интегративной деятельности третьего блока, а с другой - является одним из источников активности первого блока. Роль третьего блока проявляется в его интегрирующей, регулирующей, модулирующей функции. Обеспечивая важный этап в переработке информации, аппараты третьего блока оказывают модулирующее влияние и на первый и на второй блоки, что приводит как к изменению уровня активации коры, с одной стороны, так и к изменению порогов ощущения и восприятия - с другой.
Анализируя взаимодействие блоков мозга с точки зрения деятельности, следует отметить, что первый блок участвует в формировании мотивов любой сознательной деятельности, второй - обеспечивает операциональную сторону деятельности, а третий - отвечает за формирование целей и программ деятельности. Нарушение работы каждого из этих блоков обязательно приводит к дезинтеграции психической деятельности в целом, но каждый раз по-разному, так как приводит к нарушению соответствующих стадий деятельности.
Читайте также: