
Какие отклонения от статистических закономерностей наследования установленных менделем
Обновлено: 30.06.2024
С участием "неменделирующее наследование"Мы имеем в виду любую модель наследования, при которой унаследованные персонажи не разделяются в соответствии с законами Менделя.
- Закон разделения персонажей и
- Закон независимого распределения.
Успехи и выводы Менделя были скрыты в течение многих лет, пока они не были открыты заново в начале 20 века.
В то время, однако, научное сообщество придерживалось несколько скептической позиции по отношению к этим законам, поскольку они, казалось, не объясняли закономерностей наследственности у каких-либо видов животных или растений, особенно у тех признаков, которые определяются более чем одним локусом.
Модели неменделирующего наследования
Менделирующее наследование относится к наследственной схеме, которая соответствует законам сегрегации и независимого распределения, согласно которым ген, унаследованный от любого родителя, секретируется в гаметах с эквивалентной частотой или, лучше сказать, с такой же вероятностью.
Основными паттернами менделевского наследования, описанными для некоторых заболеваний, являются: аутосомно-рецессивный, аутосомно-доминантный и сцепленный с Х-хромосомой, которые добавляются к паттернам доминирования и рецессивности, описанным Менделем.
Однако они постулировались относительно видимых признаков, а не генов (следует отметить, что некоторые аллели могут кодировать признаки, которые выделяются как доминантные, тогда как другие могут кодировать те же признаки, но они выделяются как рецессивные гены).
Из вышеизложенного следует, что неменделирующее наследование просто состоит из любого наследственного паттерна, который не соответствует норме, при которой ген, унаследованный от любого родителя, разделяется в клетках зародышевой линии с той же вероятностью, и они включают :
- Наследование, сцепленное с полом
Возникновение этих вариаций в наследственных паттернах может быть связано с различными взаимодействиями генов с другими клеточными компонентами, в дополнение к тому факту, что каждый из них подвержен регуляции и вариациям на любой из стадий транскрипции, сплайсинга, трансляции. , сворачивание белка, олигомеризация, транслокация и компартментализация внутри клетки и для ее экспорта.
Митохондриальное наследование
Митохондриальная ДНК также передает информацию от одного поколения к другому, как и та, что содержится в ядрах всех эукариотических клеток. Геном, закодированный в этой ДНК, включает гены, необходимые для синтеза 13 полипептидов, которые являются частью субъединиц дыхательной цепи митохондрий, необходимых для организмов с аэробным метаболизмом.
“Печать” или отпечаток гена
Существуют кластеры импринтинга генов, которые состоят из от 3 до 12 генов, распределенных между 20 и 3700 килограммами оснований ДНК. Каждый кластер имеет область, известную как область контроля импринтинга, которая демонстрирует определенные эпигенетические модификации от каждого родителя, включая:
- метилирование ДНК по определенным аллелям в остатках цитокинов пар CpG
- Посттрансляционная модификация гистонов, связанных с хроматином (метилирование, ацетилирование, фосфорилирование и т. Д. Аминокислотных хвостов этих белков).
Паттерны наследования, в которых проявление заболевания зависит от конкретных аллелей, унаследованных от любого из родителей, известны как эффект родительского происхождения.
Однородительская дисомия
Это явление является исключением из первого закона Менделя, который гласит, что только один из двух аллелей, присутствующих у каждого родителя, передается потомству и, согласно хромосомным законам наследования, может передаваться только одна из родительских гомологичных хромосом. следующему поколению.
Это исключение из правил, поскольку однопородная дисомия - это наследование обеих копий гомологичной хромосомы от одного из родителей. Этот тип наследования не всегда обнаруживает фенотипические дефекты, поскольку он сохраняет числовые и структурные характеристики диплоидных хромосом.
Неполное доминирование
Этот образец наследования состоит, фенотипически говоря, из смеси аллель-кодированных признаков, которые объединены. В случаях неполного доминирования те особи, которые являются гетерозиготными, демонстрируют смесь признаков двух контролирующих их аллелей, что означает, что отношения между фенотипами изменены.
Кодоминантность
Множественные аллели
Возможно, одна из основных слабостей менделевского наследования представлена признаками, которые кодируются более чем одним аллелем, что довольно часто встречается у людей и многих других живых существ.
Это унаследованное явление увеличивает разнообразие признаков, которые кодируются геном, и, кроме того, эти гены могут также испытывать паттерны неполного доминирования и кодоминирования в дополнение к простому или полному доминированию.
Плейотропия
Летальные аллели
В своих работах Мендель также не рассматривал наследование определенных аллелей, которые могут препятствовать выживанию потомства, когда оно находится в гомозиготной или гетерозиготной форме; это летальные аллели.
Летальные аллели обычно связаны с мутациями или дефектами генов, которые строго необходимы для выживания, которые при передаче следующему поколению (такие мутации), в зависимости от гомозиготности или гетерозиготности индивидуумов, являются летальными.
Признаки или полигенное наследование
Есть характеристики, которые контролируются более чем одним геном (с их аллелями) и, кроме того, сильно контролируются окружающей средой. У людей это чрезвычайно распространено и касается таких черт, как рост, цвет глаз, волос и кожи, а также риск заболеть некоторыми заболеваниями.
Наследование, сцепленное с полом
Большинство признаков, связанных с полом, связаны с некоторыми рецессивными заболеваниями и расстройствами.
Примеры неменделирующего наследования
У людей есть генетическое заболевание, известное как синдром Марфана, которое вызвано мутацией в одном гене, который одновременно влияет на рост и развитие (среди прочего, рост, зрение и сердечную функцию).
Это случай, который считается отличным примером неменделевского паттерна наследования, называемого плейотропией, в котором один ген контролирует несколько характеристик.
Пример митохондриального наследования
Генетические нарушения, возникающие в результате мутаций в митохондриальной ДНК, представляют собой ряд клинических фенотипических вариаций, поскольку возникает так называемая гетероплазмия, когда разные ткани имеют разный процент мутантного митохондриального генома и, следовательно, представляют разные фенотипы.
Эти синдромы могут быть связаны с мутациями в ядерном геноме, которые влияют на ядерные гены, участвующие в синтезе митохондриальных нуклеотидов или в репликации митохондриальной ДНК. Эффекты могут выражаться в миопатиях, энцефалопатиях, гепато-церебральных или нервно-желудочно-кишечных дефектах.
Гипермаркет знаний>>Биология>>Биология 9 класс>> Закономерности наследования признаков, установленные Г. Менделем
Закономерности наследования признаков, установленные Г. Менделем.
1. У каких организмов только одна кольцевая хромосома?
2. Что такое гибрид?
Генетика — наука, изучающая закономерности наследственности и изменчивости живых организмов.
Наследственность — это свойство всех живых организмов передавать свои признаки и свойства из поколения в поколение.
Изменчивость — свойство всех живых организмов приобретать в процессе индивидуального развития новые признаки. Элементарные единицы наследственности — гены — представляют собой участки ДНК хромосом.
Гибридологический метод.
Основой замечательной работы Г. Менделя был так называемый гибридологический метод. Суть этого метода заключается в скрещивании (гибридизации) организмов, отличающихся друг от друга какими-либо признаками, и в последующем анализе характера наследования этих признаков у потомства. Гибридологический метод до сих пор лежит в основе исследований всех генетиков.
Ставя опыты, Мендель придерживался нескольких правил.
Во-первых, работая с садовым горохом, он использовал лля скрещивания растения, которые относились к различным сортам. Так, например, у одного сорта горошины всегда были желтые, а у другого — всегда зеленые. Так как горох самоопыляемое растение, то в природных условиях эти сорта не смешиваются. Такие сорта называют чистыми линиями.
Во-вторых, чтобы получить больше материала для анализа законов наследственности, Мендель работал не с одной, а с несколькими родительскими парами гороха.
В-третьих, Мендель намеренно упростил задачу, наблюдая за наследованием не всех признаков гороха сразу, а только одной их пары. Для своих опытов он изначально выбрал пвет семян гороха — горошин. В тех случаях, когда родительские организмы различаются лишь по одному признаку например, только по цвету семян или только по форме семян), скрещивание называют моногибридным.
В-четвертых, имея математическое образование, Мендель применил для обработки данных количественные методы: он не просто замечал, каков цвет семян гороха у потомства, но и точно подсчитывал, сколько таких семян появилось.
Надо добавить, что Мендель очень удачно выбрал для опытов горох. Горох легко выращивать, в условиях Чехии он размножается несколько раз в год, сорта гороха отличаются пруг от друга рядом хорошо заметных признаков, и, наконец, з природе горох самоопыляем, но в эксперименте это самоопыление легко предотвратить, и экспериментатор может опылять растение пыльцой с другого растения, т. е. перекрестно.
Если пользоваться терминами, появившимися через много лет после работ Менделя, то можно сказать, что клетки растений гороха одного сорта содержат по два гена только желтой окраски, а гены растений другого сорта — по два гена только зеленой окраски. Гены, ответственные за развитие одного признака (например, цвета семян), получили название аллелъных генов. Если организм содержит два одинаковых аллельных гена (например, оба гена зеленого цвета :емян или, наоборот, оба гена желтизны семян), то такие организмы называют гомозиготными. Если же аллельные гены различны (т. е. один из них определяет желтую, а другой — зеленую окраску семян), то такие организмы называют гетерозиготными. Чистые линии образованы гомозиготными растениями, поэтому при самоопылении они всегда воспроизводят один вариант проявления признака. В опытах Менделя это был один из двух возможных цветов семян гороха — или всегда желтый, или всегда зеленый.
(Не будем забывать, что в те годы, когда Мендель ставил свои эксперименты, о генах, хромосомах, митозе и мейозе не было известно ничего!)
Единообразие гибридов первого поколения. Искусственно скрещивая растения гороха с желтыми горошинами с растениями, имеющими зеленые горошины (т. е. проводя моногибридное скрещивание), Мендель убедился, что все семена потомков-гибридов будут желтого цвета. Такое же явление он наблюдал в опыте при скрещивании растений с гладкими и морщинистыми семенами - все гибридные растения имели гладкие семена.
Проявляющийся у гибридов признак (желтизну семян или гладкость семян) Мендель назвал доминантным, а подавляемый признак (т. е. зеленый цвет семян или морщинистость семян) — рецессивным. Доминантный признак принято обозначать большой буквой (А, В, С), а рецессивный — маленькой (а, в, с).
На основании этих данных Мендель сформулировал правило единообразия гибридов первого поколения: при скрещивании двух гомозиготных организмов, отличающихся друг от друга одним признаком, все гибриды первого поколения будут иметь признак одного из родителей, и поколение по данному признаку будет единообразным.
Из семян, полученных в первом поколении, Мендель вырастил растения гороха и снова скрестил их между собой. У растений второго поколения большинство горошин были желтого цвета, но встречались и зеленые горошины. Всего от нескольких скрещиваемых пар растений Мендель получил 6022 желтых и 2001 зеленых горошин. Легко сосчитать, чтс 3/4 гибридных семян имели желтую окраску и ¼ зеленую. Явление, при котором скрещивание приводит к образованию потомства частично с доминантными, частично с рецессивными признаками, получило название расщепления.
Опыты с другими признаками подтвердили эти результаты, и Мендель сформулировал правило расщепления: при скрещивании двух потомков (гибридов) первого поколения между собой во втором поколении наблюдается расщепление и снова появляются особи с рецессивными признаками; эти особи составляют одну четвертую часть от всего числа потомков второго поколения.
Как же объяснить закономерности генетики с позиций современной науки?
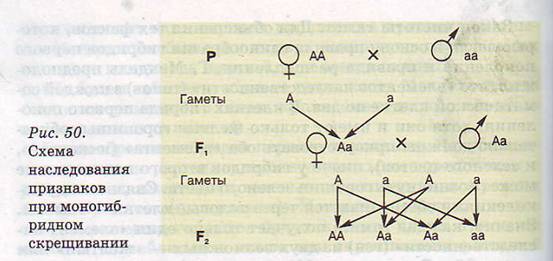
Цитологические основы закономерностей наследования при моногибридном скрещивании. Изобразим моногибридное скрещивание в виде схемы. Символ 0 обозначает женскую особь, символ 0 мужскую, х — скрещивание, Р — родительское поколение, F1— первое поколение потомков, F2— второе поколение потомков, А — ген, отвечающий за доминантный желтый цвет, а — ген, отвечающий за рецессивный зеленый цвет семян гороха (рис. 50).
Из рисунка видно, что в каждой гамете родительских особей будет по одному гену (вспомните мейоз): в одном случае А, в другом — а. Таким образом, в первом поколении все соматические клетки будут гетерозиготными — Аа. В свою счередь, гибриды первого поколения с равной вероятностью могут образовывать гаметы А или а.
Случайные комбинации этих гамет при половом процессе могут дать следующие варианты: АА, Аа, аА, аа. Первые три растения, содержащие ген А, по правилу доминирования будут иметь желтые горошины, а четвертое — рецессивная гомозигота аа —- будет иметь зеленые горошины.
Гибридологический метод. Чистые линии. Моногибридные скрещивания. Аллельные гены. Гомозиготные и гетерозиготные организмы. Доминантные и рецессивные признаки. Расщепление. Закон чистоты гамет.
1. Каких правил придерживался Г. Мендель при проведении своих опытов?
2. Почему для опытов Г. Менделя был удачным выбор гороха?
3. Какие гены называются аллельными?
4. Чем гомозиготный организм отличается от гетерозиготного?
5. В чем суть гибридологического метода?
6. Сформулируйте закон чистоты гамет.
7. Что такое моногибридное скрещивание?
8. Какой признак называется доминантным? рецессивным?
9. В чем суть правила единообразия гибридов первого поколения? Проиллюстрируйте своё ответ схемой.
10. Сформулируйте правило расщепления. Нарисуйте схему скрещивания гибридов первого поколения.
Онлайн библиотека с учениками и книгами, плани-конспекти уроков с Биологии 9 класса, книги и учебники согласно календарного плана планирование Биологии 9 класса
Если у вас есть исправления или предложения к данному уроку, напишите нам.
Если вы хотите увидеть другие корректировки и пожелания к урокам, смотрите здесь - Образовательный форум.
Отклонения наследования признаков от статистических закономерностей
Многочисленные опыты, изучающие наследование признаков у различных организмов, в большинстве своем подтвердили действенность законов Менделя. В то же время ряд факторов свидетельствовал о встречающихся отклонениях от менделевских законов. Особенно это касалось третьего закона Менделя – о независимом наследовании признаков. Оказалось, что, в отличие от первого и второго законов, которые практически универсальны, третий справедлив лишь тогда, когда исследуемые аллели генов расположены в хромосомах из разных пар (то есть в негомологичных хромосомах).
А как же наследуются гены, расположенные в одной паре гомологичных хромосом?
Большую работу по изучению наследования неаллельных генов, расположенных в парах гомологичных хромосом, осуществили американский ученый Томас Хант Морган и сотрудники его лаборатории А. Стертевант, К. Бриджес и Г.Д. Меллер в экспериментах с дрозофилой. Анализируя наблюдаемые отклонения, они пришли к выводу, что гены, расположенные в одной хромосоме, наследуются совместно – сцеплено. Это явление впоследствии получило название закона Моргана.
Совместное наследование генов, ограничивающее их свободное комбинирование, Морган назвал сцеплением генов или сцепленным наследованием.
Группа генов, для которых свойственно совместное наследование, составляет группу сцепления. Сцепленные гены расположены в хромосомах в линейном порядке. Число групп сцепления соответствует числу пар хромосом, то есть гаплоидному набору, и постоянно для каждого вила организмов. Например, у человека 46 хромосом и 23 группы сцепления, у дрозофилы 8 хромосом и 4 группы сцепления.
Гены, входящие в одну группу сцепления, не подчиняются закону независимого наследования. При дигибридном скрещивании расщепление по фенотипу происходит в соотношении не 9 : 3 : 3 : 1, а 3 : 1 (как при моногибридном скрещивании).
Линейный порядок расположения генов характерен для групп сцепления у всех организмов, включая человека.
Сцепление генов приводит к более низкой вероятности появления особей с иными (неродительскими) сочетаниями признаков, чем ожидается при их независимом наследовании.
Изучение сцепления генов и сцепленного наследования признаков послужило началом разработки теории кроссинговера (от англ. crossingover – перекрест).
Явление кроссинговера
Исследования генетиков показали, что аллели одного гена в гомологичной хромосоме могут меняться местами и перемещаться из одной гомологичной хромосомы в другую – парную. Такой обмен генами в гомологичных парах хромосом происходит регулярно. Процесс обмена идентичными участками гомологичных хромосом с содержащимися в них генами называют кроссинговером или перекрестом хромосом. Кроссинговер приводит к новым комбинациям аллелей разных генов путем взаимного обмена гомологичными участками гомологичных хромосом в результате разрыва в их нитях (хроматидах) и соединения образовавшихся фрагментов в новой последовательности. Места разрыва и соединения нитей хромосомы называют хиазмами. Перекресты в хромосомах бывают одинарные и множественные.
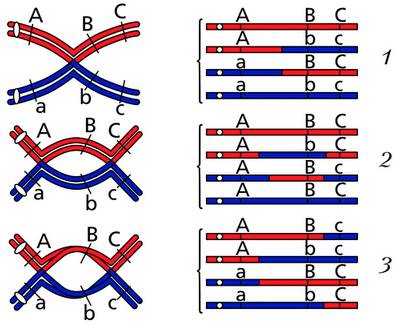
Кроссинговер (A, B, C и a, b, c – обозначения генов): 1 – одиночная хиазма; 2 – двухцепочечные двойные хиазмы; 3 – четырехцепочечные двойные хиазмы
Частота встречаемости перекрестов между генами отражает расстояние между этими генами в хромосоме. Чем больше расстояние, тем меньше сила сцепления между ними и, следовательно, чаще образуются путем кроссинговера рекомбинантные (преобразованные) типы гамет. И наоборот, чем меньше расстояние, тем сильнее сцепление и, следовательно, чаще образуются гаметы родительского типа.
Частота кроссинговера хромосом отражает силу сцепления генов в хромосоме: чем она больше, тем меньше сила сцепления.
Различают неполное и полное сцепление генов. Обычно происходит неполное сцепление генов. Оно проявляется при возможности кроссинговера между генами, относящимися к одной группе сцепления. Полное сцепление возможно лишь в случаях, когда между генами, относящимися к одной группе сцепления, рекомбинация невозможна. Отсутствие кроссинговера между генами (полное сцепление) бывает очень редко, как исключение, и известно лишь у самцов дрозофилы и шелкопряда.
На основе изучения сцепления генов и кроссинговера генетики строят генетические карты хромосом, где цифрами обозначены расстояния между генами. Генетической картой хромосомы называют схему взаимного расположения генов.
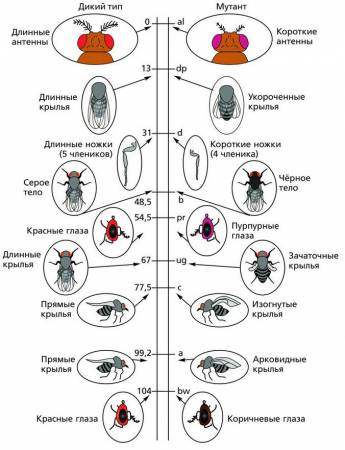
Генетические карты хромосом дрозофилы. Цифры указывают расстояние между генами и одним из концов хромосомы (в единицах перекреста)
Изучение сцепления генов и сцепленного наследования признаков послужило мощным толчком для развития исследования роли кроссинговера и явилось одним из важных подтверждений хромосомной теории наследственности.
Хромосомная теория наследственности
К началу ХХ века благодаря исследованиям генетиков был накоплен достаточно большой объем фактов и обобщений о наследственности и передаче наследственной информации от родителей к потомкам, о свойствах хромосом и отдельных генов. Все это легло в основу теории гена и хромосомной теории наследственности, разработанных Т. Морганом и сотрудниками его лаборатории. Согласно этим теориям ген является основным и элементарным носителем наследственной информации, а наследственность представляется как свойство организмов обеспечивать материальную и функциональную преемственность между поколениями и, следовательно, непрерывность жизни при смене поколений.
В 50-х годах ХХ века хромосомная теория наследственности была подкреплена исследованиями биохимиков и цитологов. Была доказана роль молекул нуклеиновых кислот, в частности ДНК, в хранении, реализации и передаче наследственной информации, показано, что молекулы ДНК являются цитологической основой генов. Расшифровка пространственной структуры молекулы ДНК, осуществленная в 1953 году, подтвердила участие этого уникального химического соединения в передаче наследственной информации.
В настоящее время установлены многие закономерности наследования свойств (признаков) организмов. Все они находят отражение в хромосомной теории наследственности. Назовем основные положения этой теории.
- Гены являются носителями наследственных свойств организмов, поэтому считаются единицами наследственной информации.
- Цитологической основой генов являются группы лежащих рядом нуклеотидов в цепочках ДНК.
- Гены, находящиеся в хромосомах ядра клетки, наследуются как отдельные независимые единицы.
- У всех организмов одного и того же вида каждый ген всегда расположен в одном и том же месте (локусе) определенной хромосомы.
- Все разновидности гена представляют собой аллеломорфы (то есть аллели), любые изменения гена приводят к появлению новых аллелей этого гена и, следовательно, к изменению признака.
- Все хромосомы и гены особи присутствуют в ее клетках всегда в виде пары, попавшей в зиготу от обоих родителей при оплодотворении.
- В каждой гамете может быть только по одной одинаковой (гомологичной) хромосоме и по одному гену из аллельной пары.
- Во время мейоза различные пары хромосом распределяются между гаметами независимо друг от друга и совершенно случайно, таким же образом наследуются и находящиеся в этих хромосомах гены.
- Важным источником появления новых комбинаций гена служит кроссинговер (перекрест хромосом).
- Развитие организмов происходит под контролем генов в тесной взаимосвязи с факторами окружающей их внутренней и внешней среды.
- Выявленные закономерности наследования признаков свойственны всем без исключения живым организмам с половым размножением.
Установлено, что ген служит основой для построения белков, большинство из которых обладают каталитической функцией. Поскольку именно ферменты управляют всеми химическими реакциями, в результате которых формируются признаки организма, указанная роль гена в реализации генетической информации является чрезвычайно важной. Это можно выразить следующим образом.
Ген → белок → химическая реакция → признак.
Современные представления о генотипе, фенотипе и геноме
Генотип, как отмечалось выше, представляет собой систему взаимодействующих генов организма (особи). Разные аллели гена могут приводить к одинаковым или различным проявлениям элементарных признаков – фенов. Все признаки организма проявляются в фенотипе – комплексе всех фенов организма. Однако однозначного соответствия между генотипом и фенотипом нет, так как изменения генотипа не всегда сопровождаются изменением фенотипа, а изменения фенотипа не всегда обусловлены изменениями генотипа. В фенотипе реализуются многие генотипические возможности организма, но не все. Поэтому фенотип обычно является частным случаем проявления генотипа в данных конкретных условиях среды. Реализуемые возможности фенотипических проявлений организма называют его нормой реакции.
Обычно генотип определяет пределы (размах) генетических возможностей, свойственных конкретному виду, а фенотип реализует эти возможности в признаках как норму реакции организма.
Генотип выражает совокупность генетической информации особи (организма). Совокупность всех генов вида (популяции) называют генофондом, а совокупность генов, содержащихся в гаплоидном наборе хромосом определенного вида организмов, – геномом.
Организмы обитают и развиваются в определенных условиях окружающей среды, испытывая на себе действие ее внешних факторов. Поэтому наследование признаков даже у близкородственных организмов может быть разным. Это явление различного наследования признаков особями в пределах вида называют изменчивостью. Наследственность и изменчивость присущи всему живому на нашей планете. Они обеспечивают преемственность наследственных свойств организмов и возможность приспособления к изменяющимся условиям среды, обусловливают поступательное развитие жизни и реализуют появление большого разнообразия организмов и видов в живой природе.
Чешский исследователь Грегор Мендель (1822–1884) считается основателем генетики, так как он первым, еще до того как оформилась эта наука, сформулировал основные законы наследования. Многие ученые до Менделя, в том числе выдающийся немецкий гибридизатор XVIII в. И. Кельрейтер, отмечали, что при скрещивании растений, принадлежащих к различным разновидностям, в гибридном потомстве наблюдается большая изменчивость. Однако объяснить сложное расщепление и, тем более, свести его к точным формулам никто не сумел из-за отсутствия научного метода гибридологического анализа.
Именно благодаря разработке гибридологического метода Менделю удалось избежать трудностей, запутавших более ранних исследователей. О результатах своей работы Г. Мендель доложил в 1865 г. на заседании Общества естествоиспытателей в г. Брюнна. Сама работа под названием “Опыты над растительными гибридами” была позже напечатана в “Трудах” этого общества, но не получила надлежащей оценки современников и оставалась забытой в течение 35 лет.
Будучи монахом, свои классические опыты по скрещиванию различных сортов гороха Г. Мендель проводил в монастырском саду в г. Брюнна. Он отобрал 22 сорта гороха, которые имели четкие альтернативные различия по семи признакам: семена желтые и зеленые, гладкие и морщинистые, цветки красные и белые, растения высокие и низкие и т.д. Важным условием гибридологического метода было обязательное использование в качестве родителей чистых, т.е. не расщепляющихся по изучаемым признакам форм.
Большую роль в успехе исследований Менделя сыграл удачный выбор объекта. Горох посевной — самоопылитель. Для получения гибридов первого поколения Мендель кастрировал цветки материнского растения (удалял пыльники) и производил искусственное опыление пестиков пыльцой мужского родителя. При получении гибридов второго поколения эта процедура уже была не нужна: он просто оставлял гибриды F1 самоопыляться, что делало эксперимент менее трудоемким. Растения гороха размножались исключительно половым способом, так что ни какие отклонения не могли исказить результаты опыта. И, наконец, у гороха Мендель обнаружил достаточное для анализа количество пар ярко контрастирующих (альтернативных) и легко различимых пар признаков.
Мендель начал анализ с самого простого типа скрещивания — моногибридного, при котором у родительских особей имеются различия по одной паре признаков. Первой закономерностью наследования, обнаруженной Менделем, было то, что все гибриды первого поколения имели одинаковый фенотип и наследовали признак одного из родителей. Этот признак Мендель назвал доминантным. Альтернативный ему признак другого родителя, не проявившийся у гибридов, был назван рецессивным. Открытая закономерность получила названия I закона Менделя, или закона единообразия гибридов I-го поколения. В ходе анализа второго поколения была установлена вторая закономерность: расщепление гибридов на два фенотипических класса (с доминантным признаком и с рецессивным признаком) в определенных числовых отношениях. Путем подсчета количества особей в каждом фенотипическом классе Мендель установил, что расщепление в моногибридном скрещивании соответствует формуле 3 : 1 (на три растения с доминантным признаком, одно — с рецессивным). Эта закономерность получила название II закона Менделя, или закона расщепления. Открытые закономерности проявлялись при анализе всех семи пар признаков, на основании чего автор пришел к выводу об их универсальности. При самоопылении гибридов F2 Мендель получил следующие результаты. Растения с белыми цветами давали потомство только с белыми цветками. Растения с красными цветками вели себя по-разному. Лишь третья часть их давала единообразное потомство с красными цветами. Потомство остальных расщеплялось в отношении красной и белой окраски в соотношении 3 : 1.
Ниже приведена схема наследования окраски цветков гороха, иллюстрирующая I и II законы Менделя.
Схема наследования красной и белой окраски цветков у гороха

При попытке объяснить цитологические основы открытых закономерностей Мендель сформулировал представление о дискретных наследственных задатках, содержащихся в гаметах и определяющих развитие парных альтернативных признаков. Каждая гамета несет по одному наследственному задатку, т.е. является “чистой”. После оплодотворения зигота получает два наследственных задатка (один — от матери, другой — от отца), которые не смешиваются и в дальнейшем при образовании гибридом гамет также попадают в разные гаметы. Эта гипотеза Менделя получила название правила “чистоты гамет”. От комбинации наследственных задатков в зиготе зависит то, каким признаком будет обладать гибрид. Задаток, определяющий развитие доминантного признака, Мендель обозначал заглавной буквой (А), а рецессивный — прописной (а). Сочетание АА и Аа в зиготе определяет развитие у гибрида доминантного признака. Рецессивный признак проявляется только при комбинации аа.
В 1902 г. В. Бетсон предложил обозначить явление парности признаков термином “аллеломорфизм”, а сами признаки, соответственно, “аллеломорфными”. По его же предложению, организмы, содержащие одинаковые наследственные задатки, стали называть гомозиготными, а содержащие разные задатки — гетерозиготными. Позже, термин “аллеломорфизм” был заменен более кратким термином “аллелизм” (Иогансен, 1926), а наследственные задатки (гены), отвечающие за развитие альтернативных признаков были названы “аллельными”.
Гибридологический анализ предусматривает реципрокное скрещивание родительских форм, т.е. использования одной и той же особи сначала в качестве материнского родителя (прямое скрещивание), а затем в качестве отцовского (обратное скрещивание). Если в обоих скрещиваниях получаются одинаковые результаты, соответствующие законам Менделя, то это говорит о том, что анализируемый признак определяется аутосомным геном. В противном случае имеет место сцепление признака с полом, обусловленное локализацией гена в половой хромосоме.
Схема реципрокного моногибридного скрещивания

Буквенные обозначения: Р — родительская особь, F — гибридная особь, ♀ и ♂ — женская или мужская особь (или гамета),
заглавная буква (А) — доминантный наследственный задаток (ген), строчная буква (а) — рецессивный ген.
Среди гибридов второго поколения с желтой окраской семян есть как доминантные гомозиготы, так и гетерозиготы. Для определения конкретного генотипа гибрида Мендель предложил проводить скрещивание гибрида с гомозиготной рецессивной формой. Оно получило название анализирующего. При скрещивании гетерозиготы (Аа) с линией анализатором (аа) наблюдается расщепление и по генотипу, и по фенотипу в соотношении 1 : 1.
Схема анализирующего скрещивания
Если гомозиготной рецессивной формой является один из родителей, то анализирующее скрещивание одновременно становится беккроссом — возвратным скрещиванием гибрида с родительской формой. Потомство от такого скрещивания обозначают Fb.
Закономерности, обнаруженные Менделем при анализе моногибридного скрещивания, проявлялись также и в дигибридном скрещивании, в котором родители различались по двум парам альтернативных признаков (например, желтая и зеленая окраска семян, гладкая и морщинистая форма). Однако количество фенотипических классов в F2 возрастало вдвое, а формула расщепления по фенотипу была 9 : 3 : 3 : 1 (на 9 особей с двумя доминантными признаками, по три особи — с одним доминантным и одним рецессивным признаком и одна особь с двумя рецессивными признаками).
Для облегчения анализа расщепления в F2 английский генетик Р. Пеннет предложил его графическое изображение в виде решетки, которую стали называть по его имени (решеткой Пеннета). Слева по вертикали в ней располагаются женские гаметы гибрида F1, справа — мужские. Во внутренние квадраты решетки вписываются сочетания генов, возникающие при их слиянии, и соответствующий каждому генотипу фенотип. Если гаметы располагать в решетке в той последовательности, какая представлена на схеме, то в решетке можно заметить порядок в расположении генотипов: по одной диагонали располагаются все гомозиготы, по другой — гетерозиготы по двум генам (дигетерозиготы). Все остальные клетки заняты моногетерозиготами (гетерозиготами по одному гену).
Расщепление в F2 можно представить, используя фенотипические радикалы, т.е. указывая не весь генотип, а только гены, которые определяют фенотип. Эта запись выглядит следующим образом:

Черточки в радикалах означают, что вторые аллельные гены могут быть как доминантными, так и рецессивными, фенотип при этом будет одинаковым.
Схема дигибридного скрещивания
(решетка Пеннета)

Общее количество генотипов F2 в решетке Пеннета — 16, но разных — 9, так как некоторые генотипы повторяются. Частота разных генотипов описывается правилом:
В F2 дигибридного скрещивания все гомозиготы встречаются один раз, моногетерозиготы — два раза и дигетерозиготы — четыре раза. В решетке Пеннета представлены 4 гомозиготы, 8 моногетерозигот и 4 дигетерозиготы.
Расщепление по генотипу соответствует следующей формуле:
1ААВВ : 2ААВb : 1ААbb : 2АаВВ : 4АаВb : 2Ааbb : 1ааВВ : 2ааВb : 1ааbb.
Сокращенно - 1 : 2 : 1 : 2 : 4 : 2 : 1 : 2 : 1.
Среди гибридов F2 только два генотипа повторяют генотипы родительских форм: ААВВ и ааbb; в остальных произошла перекомбинация родительских генов. Она привела к появлению двух новых фенотипических классов: желтых морщинистых семян и зеленых гладких.
Проведя анализ результатов дигибридного скрещивания по каждой паре признаков отдельно, Мендель установил третью закономерность: независимый характер наследования разных пар признаков (III закон Менделя). Независимость выражается в том, что расщепление по каждой паре признаков соответствует формуле моногибридного скрещивания 3 : 1. Таким образом, дигибридное скрещивание можно представить как два одновременно идущих моногибридных.
Как было установлено позже, независимый тип наследования обусловлен локализацией генов в разных парах гомологичных хромосом. Цитологическую основу менделевского расщепления составляет поведение хромосом в процессе клеточного деления и последующее слияние гамет во время оплодотворения. В профазе I редукционного деления мейоза гомологичные хромосомы коньюгируют, а затем в анафазе I расходятся к разным полюсам, благодаря чему аллельные гены не могут попасть в одну гамету. Негомологичные хромосомы при расхождении свободно комбинируются друг с другом и отходят к полюсам в разных сочетаниях. Этим обусловлена генетическая неоднородность половых клеток, а после их слияния в процессе оплодотворения — генетическая неоднородность зигот, и как следствие, генотипическое и фенотипическое разнообразие потомства.
Независимое наследование разных пар признаков позволяет легко рассчитывать формулы расщепления в ди- и полигибридных скрещиваниях, так как в их основе лежат простые формулы моногибридного скрещивания. При расчете используется закон вероятности (вероятность встречаемости двух и более явлений одновременно равна произведению их вероятностей). Дигибридное скрещивание можно разложить на два, тригибридное — на три независимых моногибридных скрещивания, в каждом из которых вероятность проявления двух разных признаков в F2 равна 3 : 1. Следовательно, формула расщепления по фенотипу в F2 дигибридного скрещивания будет:
(3 : 1) 2 = 9 : 3 : 3 : 1,
тригибридного (3 : 1) 3 = 27 : 9 : 9 : 9 : 3 : 3 : 3 : 1 и т.д.
Число фенотипов в F2 полигибридного скрещивания равно 2 n , где n — число пар признаков, по которым различаются родительские особи.
Формулы расчета других характеристик гибридов представлены в таблице 1.
Таблица 1. Количественные закономерности расщепленияв гибридном потомстве
при различных типах скрещиваний
Расщепление по фенотипу в F2
Проявление закономерностей наследования, открытых Менделем, возможно только при определенных условиях (не зависящих от экспериментатора). Ими являются:
- Равновероятное образование гибридом всех сортов гамет.
- Всевозможное сочетание гамет в процессе оплодотворения.
- Одинаковая жизнеспособность всех сортов зигот.
Если эти условия не реализуются, то характер расщепления в гибридном потомстве изменяется.
Первое условие может быть нарушено по причине нежизнеспособности того или иного типа гамет, возможной вследствие различных причин, например, негативного действия другого гена, проявляющегося на гаметическом уровне.
Второе условие нарушается в случае селективного оплодотворения, при котором наблюдается предпочтительное слияние определенных сортов гамет. При этом гамета с одним и тем же геном может вести себя в процессе оплодотворения по-разному, в зависимости от того является ли она женской или мужской.
Третье условие обычно нарушается, если доминантный ген имеет в гомозиготном состоянии летальный эффект. В этом случае в F2 моногибридного скрещивания в результате гибели доминантных гомозигот АА вместо расщепления 3 : 1 наблюдается расщепление 2 : 1. Примером таких генов являются: ген платиновой окраски меха у лисиц, ген серой окраски шерсти у овец породы ширази. (Подробнее в следующей лекции.)
Причиной отклонения от менделевских формул расщепления может также стать неполное проявление признака. Степень проявления действия генов в фенотипе обозначается термином экспрессивность. У некоторых генов она является нестабильной и сильно зависит от внешних условий. Примером может служить рецессивный ген черной окраски тела у дрозофилы (мутация ebony), экспрессивность которого зависит от температуры, вследствие чего особи гетерозиготные по этому гену могут иметь темную окраску.
Открытие Менделем законов наследования более чем на три десятилетия опередило развитие генетики. Опубликованный автором труд “Опыт работы с растительными гибридами” не был понят и по достоинству оценен современниками, в том числе Ч. Дарвиным. Основная причина этого заключается в том, что к моменту публикации работы Менделя еще не были открыты хромосомы и не был описан процесс деления клеток, составляющий, как было сказано выше, цитологическую основу менделевских закономерностей. Кроме того, сам Мендель усомнился в универсальности открытых им закономерностей, когда по совету К. Негели стал проверять полученные результаты на другом объекте — ястребинке. Не зная о том, что ястребинка размножается партеногенетически и, следовательно, у нее нельзя получить гибридов, Мендель был совершенно обескуражен итогами опытов, никак не вписывавшимися в рамки его законов. Под влиянием неудачи он забросил свои исследования.
Признание пришло к Менделю в самом начале ХХ в., когда в 1900 г. три исследователя — Г. де Фриз, К. Корренс и Э. Чермак — независимо друг от друга опубликовали результаты своих исследований, воспроизводящих эксперименты Менделя, и подтвердили правильность его выводов. Поскольку к этому времени был полностью описан митоз, почти полностью мейоз (его полное описание завершилось в 1905 г.), а также процесс оплодотворения, ученые смогли связать поведение менделевских наследственных факторов с поведением хромосом в процессе клеточного деления. Переоткрытие законов Менделя и стало отправной точкой для развития генетики.
Первое десятилетие ХХ в. стало периодом триумфального шествия менделизма. Закономерности, открытые Менделем, были подтверждены при изучении различных признаков как на растительных, так и на животных объектах. Возникло представление об универсальности законов Менделя. Вместе с тем стали накапливаться факты, которые не укладывались в рамки этих законов. Но именно гибридологический метод позволил выяснить природу этих отклонений и подтвердить правильность выводов Менделя.
Все пары признаков, которые были использованы Менделем, наследовались по типу полного доминирования. В этом случае рецессивный ген в гетерозиготе не действует, и фенотип гетерозиготы определяется исключительно доминантным геном. Однако большое число признаков у растений и животных наследуются по типу неполного доминирования. В этом случае гибрид F1 полностью не воспроизводит признак того или другого родителя. Выражение признака является промежуточным, с большим или меньшим уклонением в ту или другую сторону.
Примером неполного доминирования может быть промежуточная розовая окраска цветков у гибридов ночной красавицы, полученных при скрещивании растений с доминантной красной и рецессивной белой окраской (см. схему).
Схема неполного доминирования при наследовании окраски цветков у ночной красавицы

Как видно из схемы, в скрещивании действует закон единообразия гибридов первого поколения. Все гибриды имеют одинаковую окраску — розовую — в результате неполного доминирования гена А. Во втором поколении разные генотипы имеют ту же частоту, что и в опыте Менделя, а изменяется только формула расщепления по фенотипу. Она совпадает с формулой расщепления по генотипу — 1 : 2 : 1, так как каждому генотипу соответствует свой признак. Это обстоятельство облегчает проведение анализа, так как отпадает надобность в анализирующем скрещивании.
Существует еще один тип поведения аллельных генов в гетерозиготе. Он называется кодоминированием и описан при изучении наследования групп крови у человека и ряда домашних животных. В этом случае у гибрида, в генотипе которого присутствуют оба аллельных гена, в равной мере проявляются оба альтернативных признака. Кодоминирование наблюдается при наследовании групп крови системы А, В, 0 у человека. У людей с группой АВ (IV группа) в крови присутствуют два разных антигена, синтез которых контролируется двумя аллельными генами.
Похожие материалы по теме "Законы Менделя":
Перейти к чтению других тем книги "Генетика и селекция. Теория. Задания. Ответы":
Читайте также: