
Как осуществляется процесс трансляции расскажите о роли ррнк в обеспечении
Обновлено: 03.07.2024
\u0412 \u0441\u0430\u043c\u043e\u043c \u043d\u0430\u0447\u0430\u043b\u0435 \u0441\u043f\u0438\u0440\u0430\u043b\u044c \u0414\u041d\u041a \u0440\u0430\u0441\u043f\u043b\u0435\u0442\u0430\u0435\u0442\u0441\u044f \u043d\u0430 \u0434\u0432\u0435 \u043d\u0438\u0442\u0438 \u043f\u043e\u0434 \u0434\u0435\u0439\u0441\u0442\u0432\u0438\u0435\u043c \u0444\u0435\u0440\u043c\u0435\u043d\u0442\u0430, \u043a\u043e\u0442\u043e\u0440\u044b\u0439 \u043d\u0430\u0437\u044b\u0432\u0430\u0435\u0442\u0441\u044f \u0420\u041d\u041a \u2013 \u043f\u043e\u043b\u0438\u043c\u0435\u0440\u0430\u0437\u0430. \u041d\u0430 \u043e\u0434\u043d\u043e\u0439 \u0438\u0437 \u0446\u0435\u043f\u043e\u0447\u0435\u043a \u0414\u041d\u041a \u043d\u0430\u0447\u0438\u043d\u0430\u0435\u0442\u0441\u044f \u0441\u0438\u043d\u0442\u0435\u0437 \u043f\u043e \u043c\u0435\u0442\u043e\u0434\u0443 \u043a\u043e\u043c\u043f\u043b\u0435\u043c\u0435\u043d\u0442\u0430\u0440\u043d\u043e\u0441\u0442\u0438 (\u0434\u043e\u0441\u0442\u0440\u0430\u0438\u0432\u0430\u043d\u0438\u0435 \u043d\u0435\u0434\u043e\u0441\u0442\u0430\u044e\u0449\u0438\u0445 \u0437\u0432\u0435\u043d\u044c\u0435\u0432) \u0446\u0435\u043f\u043e\u0447\u043a\u0430 \u0420\u041d\u041a, \u043d\u043e \u0442\u043e\u043b\u044c\u043a\u043e \u0432\u043c\u0435\u0441\u0442\u043e \u0422\u0438\u043c\u0438\u043d\u0430 \u0441\u0442\u0440\u043e\u0438\u0442\u0441\u044f \u0443\u0440\u0430\u0446\u0438\u043b. \u0410\u2026.. \u0422 \u0410
\u0413 \u2026..\u0426 \u0413
\u0426 \u2026..\u0413 \u0426
\u0422 \u2026..\u0410 \u0423
\u0422 \u2026..\u0410 \u0423
\u0410 \u2026..\u0422 \u0410
\u0418 \u0442. \u0434.
\u041a\u043e\u0433\u0434\u0430 \u0441\u0438\u043d\u0442\u0435\u0437 \u0437\u0430\u0432\u0435\u0440\u0448\u0430\u0435\u0442\u0441\u044f (\u0442. \u0435. \u0437\u0430\u0432\u0435\u0440\u0448\u0430\u0435\u0442\u0441\u044f \u043f\u0440\u043e\u0446\u0435\u0441\u0441 \u0441\u0447\u0438\u0442\u044b\u0432\u0430\u043d\u0438\u044f \u0438\u043d\u0444\u043e\u0440\u043c\u0430\u0446\u0438\u0438) , \u0414\u041d\u041a \u043e\u043f\u044f\u0442\u044c \u0437\u0430\u043a\u0440\u0443\u0447\u0438\u0432\u0430\u0435\u0442\u0441\u044f \u0432 \u0441\u043f\u0438\u0440\u0430\u043b\u044c \u0438 \u043e\u043d\u0430 \u043d\u0430\u043c \u0431\u043e\u043b\u044c\u0448\u0435 \u043d\u0435 \u043d\u0443\u0436\u043d\u0430, \u0432\u0435\u0434\u044c \u0435\u0435 \u043c\u0430\u0442\u0440\u0438\u0446\u0443 \u043c\u044b \u0443\u0436\u0435 \u043f\u043e\u043b\u0443\u0447\u0438\u043b\u0438 \u043d\u0430 \u0438 \u2013 \u0420\u041d\u041a. \u0418 - \u0420\u041d\u041a \u0432\u044b\u0445\u043e\u0434\u0438\u0442 \u0438\u0437 \u044f\u0434\u0440\u0430 \u0432 \u0446\u0438\u0442\u043e\u043f\u043b\u0430\u0437\u043c\u0443 \u0433\u0434\u0435 \u0438 \u043f\u0440\u043e\u0438\u0441\u0445\u043e\u0434\u0438 \u0441\u0432\u044f\u0437\u044c \u0441 \u0440\u0438\u0431\u043e\u0441\u043e\u043c\u043e\u0439.
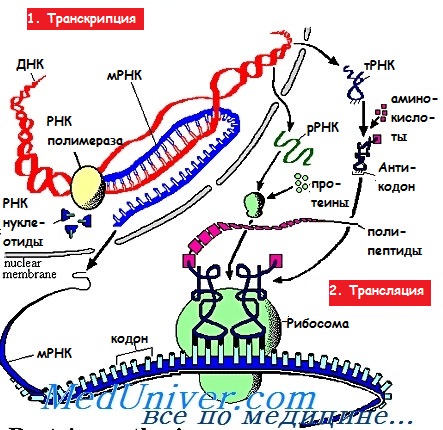
\u0422\u0440\u0430\u043d\u0441\u043b\u044f\u0446\u0438\u044f \u2013 \u044d\u0442\u043e \u043f\u0440\u043e\u0446\u0435\u0441\u0441 \u043f\u0435\u0440\u0435\u0432\u043e\u0434\u0430 \u043a\u043e\u0434\u043e\u0432\u043e\u0439 \u043f\u043e\u0441\u043b\u0435\u0434\u043e\u0432\u0430\u0442\u0435\u043b\u044c\u043d\u043e\u0441\u0442\u0438 \u043d\u0443\u043a\u043b\u0435\u043e\u0442\u0438\u0434\u043e\u0432 \u0438 \u0420\u041d\u041a \u0432 \u043f\u0435\u0440\u0432\u0438\u0447\u043d\u0443\u044e \u0441\u0442\u0440\u0443\u043a\u0442\u0443\u0440\u0443 \u0431\u0435\u043b\u043a\u0430.
Транскрипция это переписывание информации.
В самом начале спираль ДНК расплетается на две нити под действием фермента, который называется РНК – полимераза. На одной из цепочек ДНК начинается синтез по методу комплементарности (достраивание недостающих звеньев) цепочка РНК, но только вместо Тимина строится урацил. А….. Т А
Г …..Ц Г
Ц …..Г Ц
Т …..А У
Т …..А У
А …..Т А
И т. д.
Когда синтез завершается (т. е. завершается процесс считывания информации) , ДНК опять закручивается в спираль и она нам больше не нужна, ведь ее матрицу мы уже получили на и – РНК. И - РНК выходит из ядра в цитоплазму где и происходи связь с рибосомой.
Трансляция – это процесс перевода кодовой последовательности нуклеотидов и РНК в первичную структуру белка.
В цитоплазме возле ядра и-РНК поджидает т-РНК, которая зацепляет ее и ведет к рибосомам.
У транспортной РНК очень сложное строение. Она во-первых специфична, т. е. у каждой молекулы т-РНК имеется своя аминокислота, которая определяется строением триплета т-РНК. Она имеет сложную форму строения. И если схематично нарисовать, то виде креста. На одной стороне находится антикодон – триплет, а на противоположной – аминокислота, которой он соответствует.
На рибосомах происходит построение белка. Информационная РНК как бы протягивается между телом рибосомы - рис 15. А к ней подходит т-РНК. Если по методу комплементарности происходит состыковка антикодона с и-РНК, то аминокислота отходит от т-РНК и отправляется в цепочку белка, а если нет, то т-РНК ее ведет обратно. Считав информацию с одного триплета, переход на следующий. Пока вся цепочка белка не будет построена. f
Каждая клетка содержит тысячи белков. Свойства белков зависят от их первичной структуры , т. е. порядка соединения аминокислотных остатков в молекулах.
Информация о первичной структуре всех белков организма закодирована последовательностью нуклеотидов, образующих молекулы ДНК. В молекулах ДНК выделяют гены . Каждый ген соответствует одному белку.
Ген — это единица наследственности, представляющая собой участок ДНК, в котором закодирована первичная структура молекул одного белка.

Биосинтез происходит в клетках с огромной скоростью. В организме высших животных в одну минуту образуется до \(60\) тыс. пептидных связей.
Транскрипция — это процесс переписывания наследственной информации с молекулы ДНК на информационную (матричную) РНК.

Информационная (матричная) РНК одноцепочечная, она собирается на одной из нитей ДНК по правилу комплементарности.
Образуется молекула иРНК, которая является копией второй цепочки ДНК, только в ней тимин заменён на урацил. Закодированная в ДНК информация о первичной структуре белка таким образом переписывается на иРНК.
Молекула ДНК содержит большое количество генов. В начале каждого гена располагается промотор — особая последовательность нуклеотидов ДНК, которую определяет РНК-полимераза, и с этого места начинает сборку молекулы иРНК.
В клетках прокариот иРНК образуется в цитоплазме, поэтому образовавшиеся молекулы могут сразу принимать участие в синтезе белков на рибосомах.
В клетках эукариот транскрипция происходит в ядре, поэтому иРНК сначала через поры в ядерной мембране выходит в цитоплазму.
Для сборки белковой молекулы в цитоплазме клетки должны присутствовать все необходимые аминокислоты. Они образуются при расщеплении белков, поступающих с пищей, или синтезируются в самом организме.
Аминокислоты доставляются к рибосомам транспортными РНК (тРНК). Аминокислота попадает в рибосому только в комплексе с сответствующей тРНК.
К кодону, расположенному в активном центре рибосомы, присоединяется тРНК с комплементарным антикодоном. Соединённая с ней аминокислота образует пептидную связь к растущей полипептидной цепочкой. Затем рибосома перемещается на следующий кодон иРНК. В рибосоме оказывается тРНК с антикодоном, комплементарным следующему триплету в иРНК, и к образующейся молекуле белка присоединяется следующая аминокислота.

Рибосома постепенно сдвигается по иРНК, задерживаясь на следующих триплетах. Так поэтапно собирается молекула белка.
Синтез полипептидной цепи заканчивается, когда в активном центре рибосомы оказывается стоп-кодон (УАА, УАГ или УГА). Молекула белка отсоединяется от рибосомы, выходит в ЭПС или цитоплазму и усложняется, образуя характерную вторичную, третичную и четвертичную структуры.
На одной иРНК одновременно находятся несколько рибосом и происходит синтез нескольких молекул белка. Рибосомы, которые связаны с одной иРНК и синтезируют один и тот же белок, образуют полисому .
Когда синтез данного белка окончен, рибосома может найти другую иРНК и начать синтезировать другой белок.

последовательность нуклеотидов матричной цепи ДНК: ААГ ГЦТ ТАГ.
При транскрипции на этой цепи по принципу комплементарности образуется участок иРНК с нуклеотидами УУЦ ЦГА АУЦ, на котором в результате трансляции образуется цепочка из аминокислот: фенилаланин — аргинин — серин.
Если в одном из триплетов произойдёт замена нуклеотидов или они поменяются местами, то может случиться так, что триплет станет кодировать какую-нибудь другую аминокислоту. Значит, произойдут изменения и в строении белка, закодированного данным геном, что может оказать влияние на процессы обмена веществ и изменить признаки организма.
На протяжении нескольких последних десятилетий контроль над экспрессией генов на уровне трансляции стал способом регулирования роста, пролиферации, злокачественной трансформации и апоптоза. Регулирование трансляции мРНК может изменить общую трансляционную или рибосомную емкость клетки (т.е. число рибосом) либо изменить трансляционную или рибосомную эффективность (т.е. количество белка, синтезированного на одной рибосоме).
Кроме того, изменения рибосомной эффективности могут ограничиваться регулированием экспрессии конкретного белка (например, мРНК ферритина) или влиять на скорость трансляции специфических белков, которые затем участвуют в трансляции мРНК и, следовательно, оказывают глобальное всеобъемлющее влияние на производительность и эффективность трансляции мРНК (например, терминального олигопиримидина мРНК, который описан далее).
Изменение рибосомной емкости указывает на изменение содержания рибосомных белков и рибосомной РНК (рРНК) в клетке. Время, необходимое для модификации этих популяций, показывает, что изменить общий синтез белка на уровне рибосомной емкости могут лишь хронические заболевания или такие продолжительные состояния, как длительное голодание или неконтролируемый диабет.
На уровень РНК также влияет возраст. В частности, скорость общего оборота рРНК и транспортной РНК (тРНК) во всем организме у недоношенных младенцев в 3-4 раза выше, чем у взрослых, а оборот мРНК у новорожденных в 6 раз выше, чем у взрослых. Это подчеркивает тот факт, что высокий темп роста у новорожденных возможен благодаря способности организма создавать и использовать трансляционный аппарат.
Каждая рибосома состоит из 80 различных белков и 4 видов РНК (5S, 5.8S, 18S, 28S). Три вида РНК (5.8S, 18S, 28S) транскрибируются с одного гена (45S рибосомная ДНК (рДНК)) РНК-полимеразой типа I. Транскрипция РНК-полимеразы типа I усиливается фактором транскрипции UBF (фактором регуляции последующих звеньев сигнальной цепи). Состояния, усиливающие процессы анаболизма, могут сопровождаться повышенной экспрессией и/или активностью белка UBF, в результате увеличивается транскрипция 45S рДНК. Например, повышенная активность UBF способствует гипертрофии кардиомиоцитов новорожденных.
На рибосомную емкость также может оказывать влияние синтез рибосомного белка. Регуляция его синтеза происходит за счет наличия последовательности пиримидинов в 5-нетранслируемом участке гена. Этот фрагмент терминального олигопиримидина (TOP) преимущественно обнаруживается в мРНК, кодирующей рибосомные белки, факторы трансляции мРНК, такие как факторы элонгации eEFIA и eEF2, и поли-(А)-связывающий белок, участвующие в образовании рибосом.
Сокращение трансляции TOP мРНК снижает общую белковосинтетическую емкость клетки, поскольку TOP мРНК кодирует трансляционный аппарат. Ряд исследований связал воедино фосфатидилинозитол-3-киназу и сигнальный путь mTOR в трансляционной активации TOP мРНК, однако на сегодняшний день точная роль каждого пути в этом процессе полностью не определена. Питание продуктами, содержащими белок, повышает уровень трансляции TOP мРНК, в то время как диета, обедненная аминокислотами, его снижает.
Сигнальный путь, важный при регулировании трансляции ТОР мРНК и, следовательно, при биогенезе рибосом, включает протеинкиназу mTOR. mTOR образует отдельные сигнальные комплексы с дополнительными взаимодействующими белками, которые затем побуждают трансляционный аппарат адекватно реагировать на питательные вещества, инсулин/факторы роста и энергетический статус. Таким образом, путь, преобразующий сигнал mTOR, является важным механизмом, посредством которого эукариотические клетки настраивают свой потенциал биосинтеза белка в ответ на условия среды, окружающей клетку.
Нутриент-зависимый комплекс 1 mTOR, реагирующий как на гормональный, так и на аминокислотный статус, активирует сигнальный путь киназы рибосомного белка S6 (S6K1), контролирующий клеточную гипертрофию и гомеостаз глюкозы. Нутриент-зависимый комплекс 2 mTOR является важным фактором регуляции синтеза актинового цитоскелета и развития и ремоделирования тканей. Кроме того, mTOR контролирует клеточную пролиферацию, синтез рРНК, процессинг пре-рРНК и аутофагию. Таким образом, сигнальный путь mTOR контролирует широкий спектр различных событий, связанных с ростом и развитием.
Надо отметить, что mTOR и его сигнальные комплексы занимают центральное место во всех процессах, касающихся роста, и, следовательно, необходимы для роста новорожденных и анаболической реакции на прием нутриентов.
Регулирование рибосомной емкости дает организму возможность адаптироваться к условиям длительных изменений. С другой стороны, при необходимости изменений рибосомной эффективности возможно достичь без промедления, поскольку весь белковосинтетический аппарат уже присутствует. В перечень условий, которые изменяют рибосомную эффективность, входят питание, гипоксия и гормональные колебания. Изменения рибосомной эффективности предполагают регулирование на уровне трансляции мРНК.
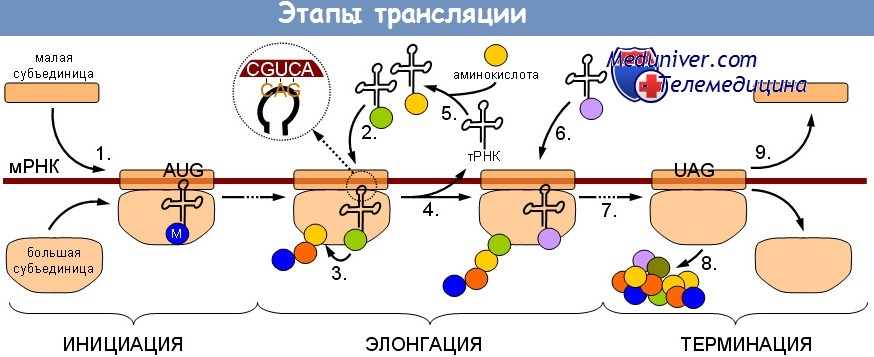
Трансляция мРНК является высокоорганизованным и многокомпонентным сигнальным путем, который можно разделить на три стадии:
(1) инициацию;
(2) элонгацию;
(3) терминацию.
На первоначальной стадии небольшие (40S) рибосомные субъединицы вовлекаются в отобранную мРНК и вместе с большой (60S) рибосомной субъединицей формируют рибосому 80S, которая способна распознать начало трансляционного кодона и начать процесс элонгации. Процесс элонгации включает энергоемкую процедуру присоединения аминокислот, одной за другой, к удлиняющейся цепочке пептида. Стадия терминации состоит из идентификации стоп-кодона и отделения рибосомных субъединиц от мРНК.
Каждая из этих стадий регулируется отдельными категориями белковых факторов, называемых соответственно стадиям эукариотическими факторами инициации (eIF), эукариотическими факторами элонгации (eEF) и эукариотическими факторами высвобождения (eRF). Большинство этих белковых факторов, регулирующих трансляцию, имеют несколько субъединиц и содержат связывающие участки для взаимодействия с другими факторами трансляции, а также для объединения с рибосомой. Кроме того, некоторые из них способны проявлять каталитическую активность, которая может быть использована для стимулирования или ингибирования трансляции.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Транскри́пция (от лат. transcriptio — переписывание) — процесс синтеза РНК с использованием ДНК в качестве матрицы, происходящий во всех живых клетках. Другими словами, это перенос генетической информации с ДНК на РНК.
Рибосо́ма — важнейшая немембранная органелла живой клетки, служащая для биосинтеза белка из аминокислот по заданной матрице на основе генетической информации, предоставляемой матричной РНК (мРНК). Этот процесс называется трансляцией. Рибосомы имеют сферическую или слегка эллипсоидную форму, диаметром от 15—20 нанометров (прокариоты) до 25—30 нанометров (эукариоты), состоят из большой и малой субъединиц.
Транспортная РНК, тРНК — рибонуклеиновая кислота, функцией которой является транспортировка аминокислот к месту синтеза белка. Имеет типичную длину от 73 до 93 нуклеотидов и размеры около 5 нм. тРНК также принимают непосредственное участие в наращивании полипептидной цепи, присоединяясь — будучи в комплексе с аминокислотой — к кодону мРНК и обеспечивая необходимую для образования новой пептидной связи конформацию комплекса.
Факторы транскрипции (транскрипционные факторы) — белки́, контролирующие процесс синтеза мРНК на матрице ДНК (транскрипцию) путём связывания со специфичными участками ДНК. Транскрипционные факторы выполняют свою функцию либо самостоятельно, либо в комплексе с другими белками. Они обеспечивают снижение (репрессоры) или повышение (активаторы) константы связывания РНК-полимеразы с регуляторными последовательностями регулируемого гена.
Упоминания в литературе
Затем готовая информационная РНК захватывается в клетке микроскопическими тельцами – рибосомами, которые сами частично построены из РНК. Рибосома перемещается вдоль информационной РНК, преодолевая за шаг один кодон, и преобразует генетический код в букву другого алфавита, состоящего из 20 разных аминокислот. Аминокислоты подносятся к месту сборки с помощью небольших молекул транспортных РНК. (Для каждой аминокислоты существует своя транспортная РНК.) По мере продвижения рибосомы вдоль информационной РНК растет цепь присоединенных аминокислот, последовательность которых точно совпадает с последовательностью соответствующих кодонов в гене. После окончания трансляции всей информационной РНК цепь аминокислот сворачивается в трехмерную структуру, форма и свойства которой полностью определяются последовательностью аминокислот. Так образуется новое химическое соединение – белок, или протеин.
Процесс трансляции очень сложен, в нем принимает участие множество действующих лиц. Главное из них – рибосома. Рибосома – это сложнейший агрегат, построенный из полусотни различных белков и молекулы РНК. Имеется в виду не та РНК, которая служит матрицей для синтеза белка на рибосоме, а другая, рибосомальная РНК (рРНК), которая является неотъемлемой частью рибосомы. Чтобы эти два класса РНК отличать друг от друга, рибосомальную РНК обозначают как рРНК, а матричную – мРНК. Рибосома – это молекулярный компьютер, переводящий тексты с нуклеотидного языка ДНК и РНК на аминокислотный язык белков. Этот узкоспециализированный компьютер работает только по одной программе, название которой – генетический код.
Два других гена ВИЧ – tat и rev – кодируются разными не связанными между собой участками генома ВИЧ (такие гены иногда называют разорванными). Они обеспечивают синтез ре-гуляторных белков, которые стимулируют транскрипцию вирусной ДНК и трансляцию РНК. Продукты этих генов способны влиять на работу не только вирусных, но и клеточных генов.
Транскрипция и процессинг происходят в ядре клетки. Затем зрелая и-РНК через поры в мембране ядра выходит в цитоплазму, и начинается трансляция.
Мы разработали единую систему понимания генетики, которая может объяснить самые разные явления, от репликации, транскрипции и трансляции генов до эволюционного происхождения видов. Однако сравнимая единая теория метаболизма, которая связала бы процессы, благодаря которым преобразования энергии и материалов, вызываемые биохимическими реакциями внутри клетки, масштабируются для поддержания жизни, обеспечения работы биологических механизмов и определения временных масштабов жизненных процессов на всех уровнях, от организмов до экосистем, формируется медленнее.
Представьте, что у вас есть кулинарная книга (геном), которая содержит множество рецептов (генов). Вы можете сделать ксерокопии отдельных рецептов и разослать их поварам. Книга у вас одна, а копий рецептов и поваров много. Такие рецепты в данной аналогии – РНК. Ну а белки – продукт деятельности поваров: различные блюда. В клетках роль поваров выполняют структуры, называющиеся рибосомами, – молекулярные фабрики для синтеза белков. Процесс синтеза белков называется трансляцией (“переводом”).
Связанные понятия (продолжение)
Промотор в генетике — последовательность нуклеотидов ДНК, узнаваемая РНК-полимеразой как стартовая площадка для начала транскрипции. Промотор играет одну из ключевых ролей в процессе инициации транскрипции.
Процессинг РНК (посттранскрипционные модификации РНК) — совокупность процессов в клетках эукариот, которые приводят к превращению первичного транскрипта в зрелую РНК.
Экспрессия генов — это процесс, в ходе которого наследственная информация от гена (последовательности нуклеотидов ДНК) преобразуется в функциональный продукт — РНК или белок. Некоторые этапы экспрессии генов могут регулироваться: это транскрипция, трансляция, сплайсинг РНК и стадия посттрансляционных модификаций белков. Процесс активации экспрессии генов короткими двуцепочечными РНК называется активацией РНК.
РНК-полимераза II — фермент эукариот, который катализирует транскрипцию ДНК, синтезирует предшественников мРНК и большинство мяРНК и микроРНК. Эта полимераза представляет собой комплекс массой 550 кДа, состоящий из 12 субъединиц. РНК-полимераза II является наиболее изученным типом РНК-полимеразы. Ей необходим широкий спектр транскрипционных факторов для того, чтобы связываться с генами выше промоторов и начинать транскрипцию.
Полиаденили́рование — это процесс присоединения большого количества остатков аденозинмонофосфата (поли(А)-хвоста) к 3'-концу первичной мРНК (пре-мРНК). Иными словами, поли(А)-хвост — это фрагмент молекулы мРНК, азотистые основания которого представлены только аденином. У эукариот полиаденилирование является частью процессинга мРНК — процесса созревания первичного транскрипта в зрелую мРНК, готовую для трансляции. Процессинг, в свою очередь, является одним из этапов экспрессии генов.
Кэп, 5'-кэп (произносится как пять-штрих-кэп), или кэп-структура (от англ. cap — шапка) — структура на 5'-конце матричных РНК (мРНК) и некоторых других РНК эукариот. Кэп состоит из одного или нескольких модифицированных нуклеотидов и характерен только для транскриптов, синтезируемых РНК-полимеразой II. Наличие кэпа — один из признаков, отличающих эукариотические мРНК от прокариотических, которые несут трифосфат на 5'-конце. Это и другие отличия обуславливают существенно более высокую стабильность.
Гисто́ны — обширный класс ядерных белков, выполняющих две основные функции: они участвуют в упаковке нитей ДНК в ядре и в эпигенетической регуляции таких ядерных процессов, как транскрипция, репликация и репарация. Существует пять различных типов гистонов H1/Н5, H2A, H2B, H3, H4. Гистоны H2A, H2B, H3, H4, называемые кóровыми гистонами (от англ. core — сердцевина), формируют нуклеосому, представляющую собой белковую глобулу, вокруг которой накручена нить ДНК. Гистон H1/H5, называемый линкерным гистоном.
Экзоны (от англ. ex(pressi)on — выражение) — участки ДНК, копии которых составляют зрелую РНК. По мнению некоторых исследователей экзоны соответствуют доменам (структурно автономным областям) в белке и являются первичными генетическими единицами, перекомбинация которых приводит к возникновению в ходе эволюции новых генов и соответственно новых белков. Экзоны чередуются в структуре гена с другими фрагментами — интронами. При альтернативном сплайсинге некоторые экзоны удаляются из зрелой РНК.
Консервати́вные после́довательности (англ. conserved sequences) — схожие или идентичные последовательности, встречающиеся в биологических полимерах: нуклеиновых кислотах, первичной и пространственной структурах белков, полисахаридах как в пределах особей разных видов (ортологичные последовательности), так и в пределах одной особи (паралогичные последовательности). Ортологичные последовательности являются подтверждением того, что определённые последовательности могут поддерживаться эволюцией, несмотря.
Комплемента́рность (в химии, молекулярной биологии и генетике) — взаимное соответствие молекул биополимеров или их фрагментов, обеспечивающее образование связей между пространственно взаимодополняющими (комплементарными) фрагментами молекул или их структурных фрагментов вследствие супрамолекулярных взаимодействий (образование водородных связей, гидрофобных взаимодействий, электростатических взаимодействий заряженных функциональных групп и т. п.).
Оперон — функциональная единица генома у прокариот, в состав которой входят цистроны (гены, единицы транскрипции), кодирующие совместно или последовательно работающие белки и объединенные под одним (или несколькими) промоторами. Такая функциональная организация позволяет эффективнее регулировать транскрипцию этих генов.
Эндонуклеазы — белки из группы нуклеаз, расщепляющие фосфодиэфирные связи в середине полинуклеотидной цепи. Эндонуклеазы рестрикции, или рестриктазы, расщепляют ДНК в определенных местах (так называемых сайтах рестрикции), они подразделяются на три типа (I, II и III) на основании механизма действия. Эти белки часто используют в генной инженерии для создания рекомбинантных ДНК, которые вводят затем в бактериальные, растительные или животные клетки.
Старт-кодон или инициаторный кодон — первый кодон матричной РНК, c которого начинается трансляция белка в рибосоме. У эукариот и архей старт-кодон всегда кодирует метионин, а у прокариот— модифицированный метионин (N-формилметионин). В большинстве случаев роль инициаторного кодона играет триплет AUG. Старт-кодону предшествует 5′-нетранслируемая область (5'-UTR). В 5'-UTR бактерий локализована последовательность Шайна — Дальгарно (AGGAGG), которая служит для связывания рибосомы и отделёна спейсером.
3′-Нетранслируемая область (3′-НТО, англ. 3′-untranslated region, 3′-UTR) — некодирующий участок мРНК, располагающийся на её 3′-конце после кодирующей области. Такое же название имеет участок ДНК, соответствующий 3′-UTR транскрипта. 3′-UTR может принимать участие в регуляции эффективности трансляции, стабильности мРНК, содержать сигналы полиаденилирования и сайты связывания микроРНК, а также выполнять ряд других регуляторных функций.
Доме́н белка́ — элемент третичной структуры белка, представляющий собой достаточно стабильную и независимую подструктуру белка, фолдинг которой проходит независимо от остальных частей. В состав домена обычно входит несколько элементов вторичной структуры. Сходные по структуре домены встречаются не только в родственных белках (например, в гемоглобинах разных животных), но и в совершенно разных белках.
Ми́кроРНК (англ. microRNA, miRNA) — малые некодирующие молекулы РНК длиной 18—25 нуклеотидов (в среднем 22), обнаруженные у растений, животных и некоторых вирусов, принимающие участие в транскрипционной и посттранскипционной регуляции экспрессии генов путём РНК-интерференции. Помимо внутриклеточной обнаружена внеклеточная (циркулирующая) микроРНК.МикроРНК кодируются ядерной ДНК растений и животных и вирусной ДНК у некоторых ДНК-содержащих вирусов.
Кле́точный цикл — период существования клетки от момента её образования путём деления материнской клетки до собственного деления или гибели.
Белковая субъединица в структурной биологии — полипептид, который вместе с другими компонентами собирается в мультимерный или олигомерный белковый комплекс. Многие природные ферменты и другие белки состоят из нескольких белковых субъединиц.
Сплайсосо́ма — структура, состоящая из молекул РНК и белков и осуществляющая удаление некодирующих последовательностей (интронов) из предшественников мРНК. Этот процесс называется сплайсингом (от англ. splicing — сращивание).
Нуклеоти́ды (нуклеозидфосфаты) — группа органических соединений, представляют собой фосфорные эфиры нуклеозидов. Свободные нуклеотиды, в частности АТФ, цАМФ, АДФ, играют важную роль в энергетических и информационных внутриклеточных процессах, а также являются составляющими частями нуклеиновых кислот и многих коферментов.
РНК-интерференция (англ. RNA interference, RNAi) — процесс подавления экспрессии гена на стадии транскрипции, трансляции, деаденилирования или деградации мРНК при помощи малых молекул РНК.
Альтернати́вный спла́йсинг — вариант сплайсинга матричных РНК (мРНК), при котором в ходе экспрессии гена на основе одного и того же первичного транскрипта (пре-мРНК) происходит образование нескольких зрелых мРНК. Структурные и функциональные различия образовавшихся транскриптов могут быть вызваны как выборочным включением в зрелую мРНК экзонов первичного транскрипта, так и сохранением в ней частей интронов. Наиболее распространённая разновидность альтернативного сплайсинга предусматривает пропуск.
Стоп-кодон или кодон терминации — единица генетического кода, тройка нуклеотидных остатков (триплет) в ДНК — кодирующая прекращение (терминацию) синтеза полипептидной цепи (трансляцию).
Деградосо́ма (англ. degradosome) — мультибелковый бактериальный комплекс, который участвует в процессинге рибосомальной РНК и деградации матричной РНК, регулируется некодирующими РНК. Он состоит из РНК-хеликазы B, рибонуклеазы Е (РНКазы Е), полинуклеотидфосфорилазы, а также гликолитического фермента енолазы. Деградосому можно изучать с помощью электронной микроскопии.
Кодо́н (кодирующий тринуклеотид) — единица генетического кода, тройка нуклеотидных остатков (триплет) в ДНК или РНК, обычно кодирующих включение одной аминокислоты. Последовательность кодонов в гене определяет последовательность аминокислот в полипептидной цепи белка, кодируемого этим геном.
Транскрипт — молекула РНК, образующаяся в результате транскрипции (экспрессии соответствующего гена или участка ДНК).
Изоформа белка — любая из нескольких разных форм одного и того же белка. Различные формы белка могут быть образованы связанными генами, или могут возникнуть из того же гена путём альтернативного сплайсинга. Большое количество изоформ вызваны однонуклеотидными полиморфизмами — небольшими генетическими различиями между аллелями одного и того же гена. Это происходит в определенных отдельных местах расположения нуклеотидов на гене.
Цис-регуляторные элементы (или цис-элементы) — участки ДНК или РНК, регулирующие экспрессию генов, находящихся на той же молекуле (обычно хромосоме).
Открытая рамка считывания (англ. Open Reading Frame, ORF) — последовательность нуклеотидов в составе ДНК или РНК, потенциально способная кодировать белок. Основным признаком наличия ORF служит отсутствие стоп-кодонов (в случае РНК — обычно UAA, UGA и UAG) на достаточно длинном участке последовательности после стартового кодона (в подавляющем большинстве случаев — AUG). Поскольку в некоторых случаях стартовый и терминирующие кодоны отличаются от канонических, а также ввиду возможности супрессии (подавления.
Фосфорилирование — процесс переноса остатка фосфорной кислоты от фосфорилирующего агента-донора к субстрату, как правило, катализируемый ферментами и ведущий к образованию сложных эфиров фосфорной кислоты.
Ко́мплекс сра́щивания экзо́нов (англ. Exon junction complex, EJC) — белковый комплекс, формирующийся на пре-мРНК в месте соединения двух экзонов, которые оказались соединёнными друг с другом в ходе сплайсинга. EJC оказывает значительное влияние на контроль качества трансляцию и локализацию сплайсированной мРНК. Считается, что комплекс соединения экзонов обеспечивает позиционно-специфичную память о произошедшем акте сплайсинга. EJC состоит из стабильного гетеротетрамерного кора, который служит платформой.
Вторичная структура — конформационное расположение главной цепи (англ. backbone) макромолекулы (например, полипептидная цепь белка или цепи нуклеиновых кислот), независимо от конформации боковых цепей или отношения к другим сегментам. В описании вторичной структуры важным является определение водородных связей, которые стабилизируют отдельные фрагменты макромолекул.
Шаперо́н (англ. chaperones) — класс белка, главная функция которого состоит в восстановлении правильной нативной третичной или четвертичной структуры белка, а также образование и диссоциация белковых комплексов.
Нуклеосома — это структурная часть хромосомы, образованная совместной упаковкой нити ДНК с гистоновыми белками H2А, H2B, H3 и H4. Последовательность нуклеосом, соединенная гистоновым белком H1, формирует нуклеофиламент, или иначе нуклеосомную нить.
Циклины — семейство белков-активаторов циклин-зависимых протеинкиназ (CDK) (англ. CDK, cyclin-dependent kinases) — ключевых ферментов, участвующих в регуляции клеточного цикла эукариот. Циклины получили своё название в связи с тем, что их внутриклеточная концентрация периодически изменяется по мере прохождения клеток через клеточный цикл, достигая максимума на его определенных стадиях.
Я́дерные спе́клы, или спе́клы, или B-снурпосо́мы, или компа́ртменты фа́кторов спла́йсинга, или доме́ны SC-35, или кла́стеры интерхромати́новых гра́нул (англ. nuclear speckles) — ядерные тельца, регулирующие сплайсинг. В ядерных спеклах находятся факторы сплайсинга и малые ядерные рибонуклеопротеины. Изменения в белковом составе и функционировании ядерных спекл приводят к изменениям в альтернативном сплайсинге пре-мРНК, поэтому считается, что ключевой функцией ядерных спекл является регуляция доступности.
Экзонуклеазы — белки из группы нуклеаз, отщепляющие концевые мононуклеотиды от полинуклеотидной цепи путём гидролиза фосфодиэфирных связей между нуклеотидами.
Биосинтез белка — это многостадийный процесс синтеза и созревания белков, протекающий в живых организмах. В биосинтезе белка выделяют два основных этапа: синтез полипептидной цепи из аминокислот, происходящий на рибосомах с участием молекул мРНК и тРНК (трансляция), и посттрансляционные модификации полипептидной цепи. Процесс биосинтеза белка требует значительных затрат энергии.
Протеасо́ма (от англ. protease — протеиназа и лат. soma — тело) — многобелковый комплекс, разрушающий ненужные или дефектные белки при помощи протеолиза (химической реакции, при которой происходит разрыв пептидных связей) до коротких пептидов (4—25 аминокислотных остатков). Эти пептиды затем могут быть расщеплены до отдельных аминокислот. Протеасомы присутствуют в клетках эукариот, архей и некоторых бактерий. В эукариотических клетках протеасомы содержатся и в ядре, и в цитоплазме. Деградация.
Читайте также: